Achillobator
It was a large, heavily-built, ground-dwelling, bipedal carnivore that would have been an active feathered predator hunting with the enlarged sickle claw on each second toe.
Measuring around 4.5–5 m (15–16 ft) long and weighing between 250–350 kg (550–770 lb), Achillobator is considered to be one of the largest dromaeosaurs, along with Austroraptor, Dakotaraptor and Utahraptor.
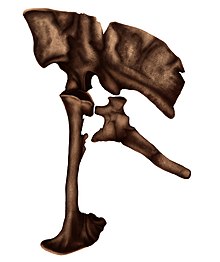
During a field exploration examining the outcrops at the Khongil locality in South Central Mongolia in 1989, conducted by the Mongolian and Russian Paleontological Expedition in the Gobi Desert, many dinosaur fossil discoveries were made.
[2] Turner and colleagues, in 2012, during their large revision of Dromaeosauridae, stated that the describing paper of Achillobator was likely published without the knowledge of the two latter paleontologists, as indicated by a draft left in Mongolia in 1997.
[4] In 2010, paleontologist Mahito Watabe and colleagues reported that additional postcranial elements remains were found, all of which belonging to a large-sized dromaeosaurid.
[5] In 2007, Mongolian paleontologist Rinchen Barsbold and team reported new dinosaur fossil findings at the Shine Us Khuduk locality of the Bayan Shireh Formation.
The above differences led Burnham and team in 2000 to suggest that the holotype of Achillobator represents a paleontological chimera and only the pedal unguals may have come from a dromaeosaurid-grade dinosaur.
[7] However, given that the specimen was actually found in semiarticulation,[8] as well as all the elements being the same color and preservation quality, the assignment of remains to a single individual is supported.
[8] Despite the fact that Achillobator features unusual and primitive characteristics compared to other dromaeosaurids, it is commonly recovered as a taxon falling within Dromaeosauridae in cladistic analyses.
[3] Achillobator represents one of the largest described dromaeosaurid taxa, reaching up to 4.5–5 m (15–16 ft) long and weighing around 250–350 kg (550–770 lb) in body mass.
[9][10][11] Although the holotype was found lacking traces of feather integument,[1] strong evidence coming from other relatives suggests the likely presence of plumage on Achillobator.
[12][13] According to the revised diagnosis by Turner and colleagues in 2012, Achillobator can be distinguished based on the following combination of characteristics and autapomorphies: The promaxillary fenestra is completely exposed; the promaxillary and maxillary fenestrae are elongated and vertically oriented at same level in the maxilla; metatarsal III is wide on the upper end; the femur is longer than the tibia; the pelvis is propubic (pubis extends forward towards the head of the animal); the obturator process on the ischium is large and triangular situated on the upper half of ischial shaft; and the boot at distal symphysis of the pubis is developed in a cranial and caudal aspect.
The neural processes of the dorsal vertebrae show resemblance to those of large ratites such as the extinct moa and extant emu, ostrich, and cassowary, and they had protruding interspinous ligaments scars that indicates a robust and similar back to those birds.
The posterior series of caudals was articulated with long chevrons and had very elongated prezygapophyses (projections of the vertebral arch that connect adjacent vertebrae).
In front and towards the back surface of the cnemial crest, however, a coarse and microsculptured area worked as the attachment for m. ilio tibialis and ambiens.
The large pubic boot had coarse areas on the dorsal surface that served as attachment sites for the m. pubo ischio femoralis internus ventralis.
[16] Eudromaeosauria was first defined as a node-based clade by Nicholas R. Longrich and Philip J. Currie in 2009 as the most inclusive natural group containing Dromaeosaurus, Velociraptor, Deinonychus, and Saurornitholestes, their most recent common ancestor and all of its other descendants.
[19] The exact position of these two vary; during the description of Halszkaraptor in 2017, Achillobator and Utahraptor were recovered as close relatives of Dromaeosaurus in the Dromaeosaurinae.
[16] The left cladogram follows Cau et al. 2017,[16] while the right cladogram follows Jasinski et al. 2020:[21] Bambiraptor Tianyuraptor Achillobator Dromaeosaurus Utahraptor Adasaurus Deinonychus Saurornitholestes Velociraptor Linheraptor Tsaagan Saurornitholestes Atrociraptor Dakotaraptor Dromaeosaurus Boreonykus Bayan Shireh dromaeosaurid Deinonychus Adasaurus Utahraptor Achillobator Acheroraptor Velociraptor Dineobellator Tsaagan Linheraptor Perle and team pointed out in 1999 that the structure of the hindlimbs and pelvic region of Achillobator indicates that the animal had massive thighs and robustly built legs suited for moderate fast-running.
[1] In 2016, Scott Persons IV and Currie examined the limb proportion of numerous theropods and found that compsognathids, troodontids and tyrannosauroids were cursorial animals with many taxa recovered with relatively high CLP (cursorial-limb-proportion) scores.
[24] Peter Mackovicky stated that the analysis might be correct on primitive dromaeosaurids (such as Microraptor) being tree-climbers, however, this does not explain why giant animals like Achillobator or Utahraptor retained sickle claws as they were likely far too large to have climbed trees with any great faculty.
For instance, most Asian species have elongated snouts based on the maxilla (animals like Velociraptor are known from complete skulls), indicating a selective feeding, such as picking up small, fast prey.
[32] Analyses on the magnetostratigraphy of the formation indicate that the entire Bayan Shireh lies within the Cretaceous Long Normal, which lasted only until the end of the Santonian stage, giving a Cenomanian-Santonian age.
[34] Over the time, a strong correlation with the Iren Dabasu Formation has been proposed by numerous authors mainly based on the similar fossil assemblages.
[37] The environments that were present in the Bayan Shireh Formation and Achillobator inhabited were relatively humid and had extensive fluvial and lacustrine facies, that is, a well-watered region dominated by prominent meanders, rivers, lakes and streams.
[28][38] Achillobator shared its surroundings in the Bayan Shireh Formation with other paleofauna such as the medium-sized theropods Erlikosaurus and Segnosaurus,[39] Garudimimus,[40] and possibly Alectrosaurus;[41] the heavy-built ankylosaurs Talarurus and Tsagantegia;[42] the small marginocephalians Amtocephale[43] and Graciliceratops;[44] the hadrosauroid Gobihadros;[32] and the large, long-necked sauropod Erketu.
[38] Non-dinosaur taxa was present as well, mostly compromising semiaquatic and terrestrial reptiles like the turtles Lindholmemys, Gobiapalone and "Trionyx";[45][46] and the crocodylomorph Paralligator.