Alternative abiogenesis scenarios
Many alternative abiogenesis scenarios have been proposed by scientists in a variety of fields from the 1950s onwards in an attempt to explain how the complex mechanisms of life could have come into existence.
[20] Bruce Damer and David Deamer argue that cell membranes cannot be formed in salty seawater, and must therefore have originated in freshwater environments like pools replenished by a combination of geothermal springs and rainfall.
[21] Experimental research at geothermal springs successfully synthesized polymers and were encapsulated in vesicles after exposure to UV light and multiple wet-dry cycles.
[23] These predecessors of true cells are assumed to have behaved more like a superorganism rather than individual structures, where the porous membranes would house molecules which would leak out and enter other protocells.
[28] Nick Lane believes that no known life forms could have utilized zinc-sulfide based photosynthesis, lightning, volcanic pyrite synthesis, or UV radiation as a source of energy.
[32] Acetyl phosphate, which is readily synthesized from thioacetate, can promote aggregation of adenosine monophosphate of up to 7 monomers which is considered energetically favored in water due to interactions between nucleobases.
Acetyl phosphate can stabilize aggregation of nucleotides in the presence of Na+ and could possibly promote polymerization at mineral surfaces or lower water activity.
[41] Reductive amination and transamination reactions catalyzed by alkaline hydrothermal vent mineral and metal ions produce amino acids.
[43] Geoffrey W. Hoffmann has argued that a complex nucleation event as the origin of life involving both polypeptides and nucleic acid is compatible with the time and space available in the primary oceans of Earth.
[45] Zachary Adam claims that tidal processes that occurred during a time when the Moon was much closer may have concentrated grains of uranium and other radioactive elements at the high-water mark on primordial beaches, where they may have been responsible for generating life's building blocks.
[46] According to computer models,[47] a deposit of such radioactive materials could show the same self-sustaining nuclear reaction as that found in the Oklo uranium ore seam in Gabon.
Such radioactive beach sand might have provided sufficient energy to generate organic molecules, such as amino acids and sugars from acetonitrile in water.
[48] In the early 1970s, Manfred Eigen and Peter Schuster examined the transient stages between the molecular chaos and a self-replicating hypercycle in a prebiotic soup.
[53] Concordantly, geochemists Szostak and Kate Adamala demonstrated that non-enzymatic RNA replication in primitive protocells is only possible in the presence of weak cation chelators like citric acid.
More abstract and theoretical arguments for metabolism without genes include Freeman Dyson's mathematical model and Stuart Kauffman's collectively autocatalytic sets in the 1980s.
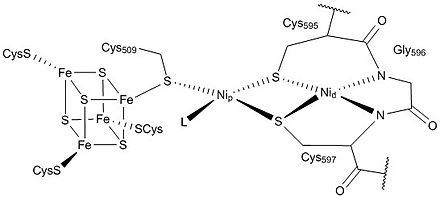
Its key enzyme unit, carbon monoxide dehydrogenase/acetyl-CoA synthase, contains mixed nickel-iron-sulfur clusters in its reaction centers and catalyzes the formation of acetyl-CoA.
[63][64] The Zn-world theory proposes that hydrothermal fluids rich in H2S interacting with cold primordial ocean (or Darwin's "warm little pond") water precipitated metal sulfide particles.
Therefore, precellular evolution may have taken place in shallow "Darwin ponds" lined with porous silicate minerals mixed with metal sulfides and enriched in K+, Zn2+, and phosphorus compounds.
[69][70] It postulates that complex organic molecules arose gradually on pre-existing, non-organic replication surfaces of silicate crystals in contact with an aqueous solution.
The clay mineral montmorillonite has been shown to catalyze the polymerization of RNA in aqueous solution from nucleotide monomers,[71] and the formation of membranes from lipids.
This system of replicating clays and their metabolic phenotype then evolved into the sulfide rich region of the hot spring acquiring the ability to fix nitrogen.
[38][75] In trying to uncover the intermediate stages of abiogenesis mentioned by Bernal, Sidney Fox in the 1950s and 1960s studied the spontaneous formation of peptide structures under plausibly early Earth conditions.
He discovered that the temperature was over 100 °C just 4 inches (100 mm) beneath the surface of the cinder cone, and suggested that this might have been the environment in which life was created—molecules could have formed and then been washed through the loose volcanic ash into the sea.
[81][82] The thermosynthesis hypothesis does not even invoke a pathway: ATP synthase's binding change mechanism resembles a physical adsorption process that yields free energy.
[83] The described first protein may be simple in the sense that it requires only a short sequence of conserved amino acid residues, a sequent sufficient for the appropriate catalytic cleft.
Sutherland has demonstrated high-yielding routes to cytidine and uridine ribonucleotides from small 2 and 3 carbon fragments such as glycolaldehyde, glyceraldehyde or glyceraldehyde-3-phosphate, cyanamide and cyanoacetylene.
[90] In 2009 they showed that the same simple building blocks allow access, via phosphate controlled nucleobase elaboration, to 2',3'-cyclic pyrimidine nucleotides directly, which can polymerize into RNA.
[99][103] A computational model (2015) has shown that virus capsids may have originated in the RNA world and served as a means of horizontal transfer between replicator communities.
[105] Tony Jia and Kuhan Chandru have proposed spontaneously-forming membraneless polyester droplets in early cellularization before the innovation of lipid vesicles.
First synthesized in 1963 from simple minerals and basic organics while exposed to sunlight, it is reported to have some metabolic capabilities, the presence of a semipermeable membrane, amino acids, phospholipids, carbohydrates and RNA-like molecules.