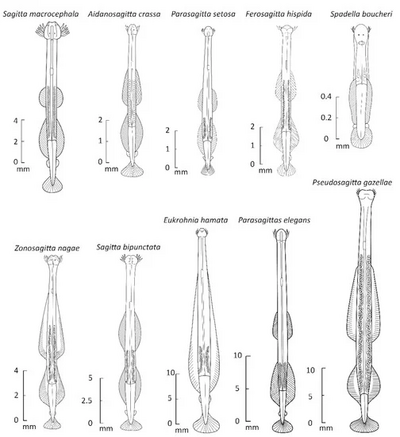
Chaetognatha
The Chaetognatha /kiːˈtɒɡnəθə/ or chaetognaths /ˈkiːtɒɡnæθs/ (meaning bristle-jaws) are a phylum of predatory marine worms that are a major component of plankton worldwide.
This organ contains wax esters which may assist reproduction and growth outside of the production season for Eukrohnia hamata in Arctic seas.
[7] Owing to the position of the oil vacuole in the center of the tractus, the organ may also have implications for buoyancy, trim and locomotion.
[3] The nervous system is reasonably simple and shows a typical protostome anatomy,[3] consisting of a ganglionated nerve ring surrounding the pharynx.
A putative sensory structure of unknown function, the retrocerebral organ, is also hosted by the posterior neuropil domain.
[3] In addition, there are a number of sensory bristles arranged in rows along the side of the body, where they probably perform a function similar to that of the lateral line in fish.
[3] A unique feature of the chaetognath eye is the lamellar structure of photoreceptor membranes, containing a grid of 35–55 nm wide circular pores.
[3] Muscles are directly excitable by electrical currents or strong K+ solutions; the main neuromuscular transmitter is acetylcholine.
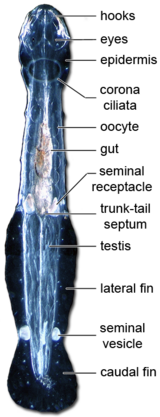
Immature sperm are released from the testes to mature inside the cavity of the tail, and then swim through a short duct to a seminal vesicle where they are packaged into a spermatophore.
The sperm rapidly escape from the spermatophore and swim along the midline of the animal until they reach a pair of small pores just in front of the tail.
Chaetognaths do not undergo metamorphosis nor they possess a well-defined larval stage,[6][3] an unusual trait among marine invertebrates;[14] however there are significant morphological differences between the newborn and the adult, with respect to proportions, chitinous structures and fin development.
They inhabit very diverse environments, from hydrothermal vents to deep ocean seafloor, to seagrass beds and marine caves.
[14] Arrow worms however engage in diel vertical migration, spending the day at lower depths to avoid predators, and coming close to the surface at night.
[3] All chaetognaths are ambush predators, preying on other planktonic animals, mostly copepods and cladocerans[6][3] but also amphipods, krill and fish larvae.
[14] Chaetognaths in the past have been traditionally, but erroneously, classed as deuterostomes by embryologists due to deuterostome-like features in the embryo.
[3] Heterokrohniidae Eukrohniidae Spadellidae Krohnittidae Sagittidae Due to their soft bodies, chaetognaths fossilize poorly.
Complete body fossils have been formally described from the Lower Cambrian Maotianshan shales of Yunnan, China (Eognathacantha ercainella Chen & Huang[41] and Protosagitta spinosa Hu[42]) and the Middle Cambrian Burgess Shale of British Columbia (Capinatator praetermissus.)
[43] A more recent chaetognath, Paucijaculum samamithion Schram, has been described from the Mazon Creek biota from the Pennsylvanian of Illinois.
It is now thought that protoconodont elements (e.g., Protohertzina anabarica Missarzhevsky, 1973), are probably grasping spines of chaetognaths rather than teeth of conodonts.
Previously chaetognaths in the Early Cambrian were only suspected from these protoconodont elements, but the more recent discoveries of body fossils have confirmed their presence then.
[45] The first known description of a chaetognath has been published by Dutch naturalist Martinus Slabber in the 1770s; he also coined the name "arrow worm".
[46][5] The zoologist Henri Marie Ducrotay de Blainville also briefly mentioned probable chaetognaths but he understood them as pelagic mollusks.
[47][5] Among the early zoologists describing arrow worms, there is Charles Darwin, who took notes about them during the voyage of the Beagle and in 1844 dedicated a paper to them.
He was also the first to propose that the genus Sagitta belonged to a separate group: «At the moment, it seems most natural to regard the Sagittas as representatives of a small group of their own that makes the transition from the real annelids (first of all the lumbricines) to the nematodes, and may not be unsuitably named Chaetognathi.»[50][5] The modern systematics of Chaetognatha begins in 1911 with Ritter-Záhony[51][18] and is later consolidated by Takasi Tokioka in 1965[52][5][18] and Robert Bieri in 1991.
[22] In 2018, reanalysis of electron microscopy photographs from the 1980s allowed scientists to identify a giant virus (Meelsvirus) infecting Adhesisagitta hispida; its site of multiplication is nuclear and the virions (length: 1.25 μm) are enveloped.
[54] In 2019, reanalysis of other previous studies has shown that structures that were taken in 1967 for bristles present on the surface of the species Spadella cephaloptera,[55] and in 2003, for bacteria infecting Paraspadella gotoi,[56] were in fact enveloped and spindle-shaped giant viruses with a cytoplasmic site of multiplication.
[57] The viral species infecting P. gotoi, whose maximum length is 3.1 μm, has been named Klothovirus casanovai (Klotho being the Greek name for one of the three Fates whose attribute was a spindle, and casanovai, in tribute to Pr J.-P. Casanova who devoted a large part of his scientific life to the study of chaetognaths).