Dictyostelid
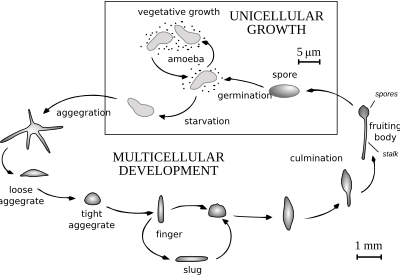
When food (normally bacteria) is readily available dictyostelids behave as individual amoebae, which feed and divide normally.
The grex has a definite anterior and posterior, responds to light and temperature gradients, and has the ability to migrate.
Under the correct circumstances the grex matures forming a sorocarp (fruiting body) with a stalk supporting one or more sori (balls of spores).
This behaviour produces beautiful spirals seen in converging colonies and is reminiscent of the Belousov–Zhabotinsky reaction and two-dimensional cyclic cellular automata.
The entire genome of Dictyostelium discoideum was published in Nature in 2005 by geneticist Ludwig Eichinger and coworkers.
Sexual development can occur when amoeboid cells are starved for their bacterial food supply and dark humid conditions are present.
As growth proceeds, the nuclei swell, and then fuse forming a true diploid zygote giant cell.
Most currently accepted phylogenies of dictyostelids utilize genome sequencing and small subunit ribosomal DNA (ssu-rDNA).
[10] Fossil calibrations indicate that dictyostelids class originally diverged into two major branches approximately 0.52 billion years ago.
[12] Cavenderia Acytostelium Heterostelium Rostrostelium Dictyostelium Polysphondylium Speleostelium Tieghemostelium Hagiwaraea Raperostelium Class Dictyostelia Lister 1909 em.
The cytoskeletal composition of D. discoideum is similar to that of mammalian cells as are the processes driven by these components, such as phagocytosis, membrane trafficking, endocytic transit and vesicle sorting.
Hence, D. discoideum represents a suitable model system to ascertain the influence of a variety of host cell factors during Legionella infections.