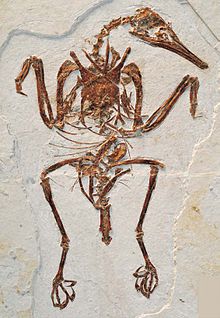
Enantiornithes
Walker made this discovery based on some partial remains from the late Cretaceous period of what is now Argentina, which he assigned to a new genus, Enantiornis, giving the entire group its name.
[7] In his paper, Walker explained what he meant by "opposite": Perhaps the most fundamental and characteristic difference between the Enantiornithes and all other birds is in the nature of the articulation between the scapula [...] and the coracoid, where the 'normal' condition is completely reversed.
For example, Alan Feduccia stated in 1996: The birds are so named because, among many distinctive features, there is a unique formation of the triosseal canal and the metatarsals are fused proximally to distally, the opposite of that in modern birds[10]Feduccia's point about the tarsometatarsus (the combined upper foot and ankle bone) is correct, but Walker did not use this reasoning in his original paper.
However, several scientists have noted that this is incorrect, because following the standard rules for forming the names of animal groups, it implies reference only to the subfamily Enantiornithinae.
[13] Birds with confidently identified characteristics of Enantiornithes found in Albian of Australia, Maastrichtian of South America, and Campanian of Mexico (Alexornis[14]), Mongolia and western edge of prehistoric Asia suggest a worldwide distribution of this group or in the relatively warm regions, at least.
Fossils attributable to this group are exclusively Cretaceous in age, and it is believed that the Enantiornithes became extinct at the same time as their non-avialan dinosaur relatives.
Almost all specimens that are complete, in full articulation, and with soft tissue preservation are known from Las Hoyas in Cuenca, Spain and the Jehol group in Liaoning (China).
Extraordinary remains of Enantiornithes have also been preserved in Burmese amber deposits dated to 99 million years ago and include hatchlings described in 2017[16] and 2018,[17] as well as isolated body parts such as wings[18][19][20] and feet.
Some Enantiornithes may have had their temporal fenestrae (holes in the side of the head) merged into the orbits as in modern birds due to the postorbitals either not being present or not being long enough to divide the openings.
[30] The presence of these primitive features of the skull would have rendered the Enantiornithes capable of only limited cranial kinesis (the ability to move the jaw independent of the cranium).
In general, the wings of Enantiornithes were advanced compared to more primitive avialans like Archaeopteryx, and displayed some features related to flight similar to those found in the lineage leading to modern birds, the Ornithuromorpha.
While most Enantiornithes had claws on at least some of their fingers, many species had shortened hands, a highly mobile shoulder joint, and proportional changes in the wing bones similar to modern birds.
Like modern birds, Enantiornithes had alulas, or "bastard wings", small forward-pointing arrangements of feathers on the first digit that granted higher maneuverability in the air and aided in precise landings.
[34] Clarke et al. (2006) surveyed all fossils of Enantiornithes then known and concluded that none had preserved tail feathers that formed a lift-generating fan, as in modern birds.
Instead of a feather fan, most Enantiornithes had a pair of long specialized pinfeathers similar to those of the extinct Confuciusornis and certain extant birds-of-paradise.
More importantly, soft tissue preserved around the tail was interpreted as the remains of a rectrical bulb, suggesting that this feature was not in fact restricted to species with modern-looking pygostyles, but might have evolved much earlier than previously thought and been present in many Enantiornithes.
This is considered at odds with the high diversity of diets that their different teeth and skull shapes imply,[44] though some modern birds have lost the gizzard and rely solely on strong stomachal acids.
[49] A 2022 study however does find them most likely to be generalistic insectivores (sans possibly Shengjingornis due to its larger size, poorly preserved skull and unusual pedal anatomy), being too small for specialised carnivory and herbivory; the atypical rostrum is tentatively speculated to be unrelated to feeding ecology.
The authors concluded that this association was a regurgitated pellet and, from the details of the digestion and the size, that the hatchlings were swallowed whole by a pterosaur or small theropod dinosaur.
[70] Evidence from nesting sites shows that Enantiornithes buried their eggs like modern megapodes, which is consistent with their inferred superprecocial adaptations.
[74] Because many Enantiornithes lacked complex tails and possessed radically different wing anatomy compared to modern birds, they have been the subject of several studies testing their flight capabilities.
[35][37][76] However, several studies have shown that they were efficient flyers, like modern birds, possessing a similarly complex nervous system and wing feather ligaments.
Enantiornithes were more advanced than Archaeopteryx, Confuciusornis, and Sapeornis, but in several respects they were more primitive than modern birds, perhaps following an intermediate evolutionary path.
This means that Enantiornithes were a successful branch of avialan evolution, but one that diversified entirely separately from the lineage leading to modern birds.
In 2010, paleontologists Jingmai O'Connor and Gareth Dyke outlined a number of criticisms against the prevailing practices of scientists failing to describe many specimens in enough detail for others to evaluate thoroughly.
Some species have been described based on specimens which are held in private collections, making further study or review of previous findings impossible.
[38] What appears fairly certain by now is that there were subdivisions within Enantiornithes possibly including some minor basal lineages in addition to the more advanced Euenantiornithes.
[87] Cruralispennia Protopteryx Elsornis Flexomornis Iberomesornis Longirostravis Rapaxavis Sinornis Enantiornis Halimornis Concornis Neuquenornis Eoalulavis Liaoningornis Gobipteryx Hebeiornis Eocathayornis Qiliania Intiornis Soroavisaurus Avisaurus Gettyia Mirarce Feitianius Longusunguis Dunhuangia Musivavis Pengornis Yuanchuavis Eopengornis Parapengornis Grabauornis Parvavis Cratoavis Gretcheniao Huoshanornis Shengjingornis Zhouornis Bohaiornis Eoenantiornis Fortunguavis Mystiornis Shenqiornis Sulcavis Key to letters: b = Boluochia c = Cathayornis e = Enantiophoenix f = Houornis h = Longipteryx i = Parabohaiornis j = Pterygornis l = Vorona m = Yuanjiawaornis n = Yungavolucris Enantiornithes taxonomy is difficult to evaluate, and as a result few clades within the group are consistently found by phylogenetic analyses.
[105] Bohaiornithids were large but geologically short-lived early Enantiornithes, with long, hooked talons and robust teeth with curved tips.
The largest and most advanced members of the group survived in North and South America up until the end of the Cretaceous, yet are very fragmentary compared to some earlier taxa.