Fish fin
It is frequently clipped off to mark hatchery-raised fish, though data from 2005 showed that trout with their adipose fin removed have an 8% higher tailbeat frequency.
[4][5] Additional information released in 2011 has suggested that the fin may be vital for the detection of, and response to, stimuli such as touch, sound and changes in pressure.
Heterocercal is the opposite of hypocercal (B) - Protocercal means the vertebrae extend to the tip of the tail and the tail is symmetrical but not expanded (as in the first fishes and the cyclostomes, and a more primitive precursor in lancelets) (C) - Homocercal where the fin usually appears superficially symmetric but in fact the vertebrae extend for a very short distance into the upper lobe of the fin.
Bony fishes (Actinopterygii and Sarcopterygii) form a taxonomic group called Osteichthyes (or Euteleostomi, which includes also land vertebrates); they have skeletons made of bone mostly, and can be contrasted with cartilaginous fishes (see below), which have skeletons made mainly of cartilage (except for their teeth, fin spines, and denticles).
Shark fin skeletons are elongated and supported with soft and unsegmented rays named ceratotrichia, filaments of elastic protein resembling the horny keratin in hair and feathers.
This is because the shark's vertebral column extends into that dorsal portion, providing a greater surface area for muscle attachment.
Similarly enlarged pectoral fins can be found in the extinct †Petalodontiformes (e.g. †Belantsea, †Janassa, †Menaspis), which belong to Holocephali (ratfish and their fossil relatives), or in †Aquilolamna (Selachimorpha) and †Squatinactis (Squatinactiformes).
In 2011, researchers using volumetric imaging techniques were able to generate "the first instantaneous three-dimensional views of wake structures as they are produced by freely swimming fishes".
For this manoeuvrability is more important than straight line speed, so coral reef fish have developed bodies which optimise their ability to dart and change direction.
[49] The pectoral and pelvic fins of many reef fish, such as butterflyfish, damselfish and angelfish, have evolved so they can act as brakes and allow complex manoeuvres.
[51] Many reef fish, such as butterflyfish, damselfish and angelfish, have evolved bodies which are deep and laterally compressed like a pancake, and will fit into fissures in rocks.
They are the posterior part of the pelvic fins that have also been modified to function as intromittent organs, and are used to channel semen into the female's cloaca during copulation.
[59][60] The oriental flying gurnard has large pectoral fins which it normally holds against its body, and expands when threatened to scare predators.
During courtship, the female cichlid, Pelvicachromis taeniatus, displays a large and visually arresting purple pelvic fin.
[69] However, recent insights from developmental patterning have prompted reconsideration of both theories in order to better elucidate the origins of paired fins.
Based on this theory, paired appendages such as pectoral and pelvic fins would have differentiated from the branchial arches and migrated posteriorly.
[72] In addition, it was later demonstrated phylogenically that pectoral and pelvic fins arise from distinct evolutionary and mechanistic origins.
[74] Although these findings do not directly support the lateral fin-fold hypothesis, the original concept of a shared median-paired fin evolutionary developmental mechanism remains relevant.
In 2009, researchers at the University of Chicago demonstrated that there are shared molecular patterning mechanisms in the early development of the chondricthyan gill arch and paired fins.
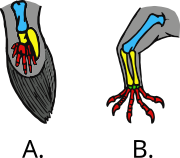
[76] In particular, terrestrial tetrapods (four-legged animals) evolved from fish and made their first forays onto land about 390 million years ago.
[79][80]Aristotle recognised the distinction between analogous and homologous structures, and made the following prophetic comparison: "Birds in a way resemble fishes.
In 2011, researchers at Monash University in Australia used primitive but still living lungfish "to trace the evolution of pelvic fin muscles to find out how the load-bearing hind limbs of the tetrapods evolved.
"[82][83] Further research at the University of Chicago found bottom-walking lungfishes had already evolved characteristics of the walking gaits of terrestrial tetrapods.
[84][85] In a classic example of convergent evolution, the pectoral limbs of pterosaurs, birds and bats further evolved along independent paths into flying wings.
Even with flying wings, there are many similarities with walking legs, and core aspects of the genetic blueprint of the pectoral fin have been retained.
Recent DNA analysis suggests that cetaceans evolved from within the even-toed ungulates, and that they share a common ancestor with the hippopotamus.
These structures are all the more remarkable because they evolved from nothing — the ancestral terrestrial reptile had no hump on its back or blade on its tail to serve as a precursor.
[98] In 2005, the Sea Life London Aquarium displayed three robotic fish created by the computer science department at the University of Essex.
"Robotic devices also facilitate three-dimensional kinematic studies and correlated hydrodynamic analyses, as the location of the locomotor surface can be known accurately.
And, individual components of a natural motion (such as outstroke vs. instroke of a flapping appendage) can be programmed separately, which is certainly difficult to achieve when working with a live animal.

(1) pectoral fins (paired), (2) pelvic fins (paired), (3) dorsal fin ,
(4) adipose fin, (5) anal fin, (6) caudal (tail) fin