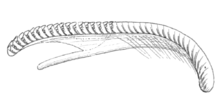
Fiona pinnata
[12] The family Fionidae was expanded in 2016 to include Tergipedidae, Eubranchidae and Calmidae as a result of a molecular phylogenetics study.
[14] Alder and Hancock's 1851 name Oithona was preoccupied by a Cyclopoid genus from W. Baird in 1843,[4][16] so in 1855 they chose instead Fiona from a character in Ossian.
[10] The oral tentacles are shorter, thickened at the base, tapering, projecting laterally and horizontally and curved backward.
The position of these glands is unusual, but there are also other species like Doto fragilis, that open into the channel of the mouth in advance of the buccal mass.
They are smooth, glossy, and of a brownish amber colour, darkest towards the anterior extremity, which gives support to the cutting blade.
This is a winglike appendage of no great size, terminating below in a free point, and having the cutting margin arched forward, plain, and nearly at right angles to the general direction of the plate.
Immediately behind this point there is the dorsal margin of the plates is reflected and expanded into an arched lobe for muscular attachment.
[18] The radula is supported on a fleshy ridge that rises up from the floor of the buccal cavity, and extends in the antero-posterior direction from the oesophagus towards the anterior opening.
The radula is long, linear, and strap-formed, and is composed of semicircular and crescent-shaped denticles (tiny teeth) of an orange colour.
The intestine leads from posterior end of the stomach, and is inclining slightly to the right side and passes backwards to the tubular anus.
The channels receive numerous branches, which communicate with the glands of the cerata, and as they approach the lateral expansion at the side of the body.
In Eolis, Embletonia, Doto, Dendronotus, Lomonotus, and Antiopa, the principal canals lie free in the visceral cavity, and in all of them there is a median posterior trunk.
In these respects Oithona would appear to resemble Hermaa, in which the whole of the hepatic ramifications are apparently connected with the skin, and there are only two principal trunks, which pass down the sides of the back.
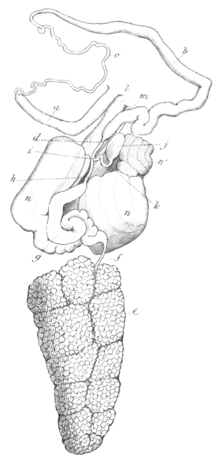
[3] On laying open the dorsal skin, the reproductive organs are found, as usual, to occupy much of the visceral cavity, having the stomach and intestine lying above, and the buccal mass in front.
[18] The ovotestis fills the posterior portion of the visceral cavity, and is composed of large irregular lobules made up almost entirely of eggs, and packed into a dense mass, tapering a little behind and truncated in front.
[verification needed] The spermoviduct leaves the anterior border of the ovary as a slender tube, but, almost immediately dilating, equals the diameter of the testis.
Spermoviduct then passes forward and suddenly contracts to its original diameter, and then advances to the anterior border of the mucous gland and receives the duct from the testis as before described.
Spermatheca is a small oval membranous sac, lying between the lobes and at the front margin of the mucous gland.
[3] The mucous gland for the secretion of the mucus-like envelope of the eggs, is composed of two lateral lobes separated on the upper surface by a deep fissure.
The heart is situated about in the middle of the back, where it forms a large oval swelling immediately below the skin, having the generative organs beneath.
The ventricle is large and muscular, of an irregular elliptical form, giving off the aorta in front, which in the usual manner supplies branches to the various organs.
The auricle is delicate in comparison with the ventricle, but is nevertheless abundantly supplied with muscular fibres; it lies diagonally in the pericardium, having the left side advanced almost to the front of that organ where it receives a trunk-vein from the skin.
Branchial ganglia form two oval central masses, resting upon the upper surface of the oesophagus, one on each side of the median line, across which they are united at the anterior extremity by a short but distinct commissure.
[3] The first pair of nerves come from the olfactory ganglions, and are large, but of no great length; they divide into several filaments as they enter the base of the dorsal tentacles.
The sixth pair are small, and come from the upper surface of the branchial ganglions; these nerves go to the skin of the sides of the back.
The seventh, much larger than the sixth, emerge from the posterior margin of the same ganglions, and supply the dorsal skin, and apparently likewise the cerata.
The posterior margins of these ganglions are united by a stout, shortish commissure, composed of two or three cords, which, passing below the gullet, form the great oesophageal collar.
The twelfth pair come from the apex of the gastro-oesophageal ganglions, and being applied to the gullet, each divides into two branches, one of which supplies the upper portion of that tube, the other, passing down it, goes to the stomach as in the other nudibranchs.
The thirteenth pair are large; these are the hepatic nerves; they issue from the buccal mass and probably[verification needed] (as in genus Aeolidia) are connected at their origin with ganglions, which must be looked upon as belonging to the sympathetic system.
The New Zealand malacologist Richard Cardeu Willan (1979)[14] published a theory that the veliger can delay its metamorphosis if it does not find suitable floating habitat to attach itself to.



{kind=link}