Heterodontosauridae
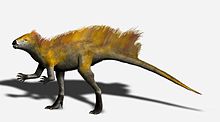
[6] Tianyulong from China appears to preserve filamentous integument which has been interpreted to be a variant of the proto-feathers found in some theropods.
In the Early Jurassic Abrictosaurus, Heterodontosaurus, and Lycorhinus, the first two premaxillary teeth were small and conical, while the much larger third tooth resembled the canines of carnivoran mammals and is often called the caniniform or 'tusk'.
The tooth rows were slightly inset from the side of the mouth, leaving a space outside the teeth that may have been bounded by a muscular cheek, which would have been necessary for chewing.
Characteristically, heterodontosaurids lacked the small openings (foramina) on the inside of the jaw bones which are thought to have aided in tooth development in most other ornithischians.
Heterodontosaurids also boasted a unique spheroidal joint between the dentaries and the predentary, allowing the lower jaws to rotate outwards as the mouth was closed, grinding the cheek teeth against each other.
The tibia and fibula of the lower leg were fused to the astragalus and calcaneum of the ankle, forming a 'tibiofibiotarsus' convergently with modern birds.
[22] Heterodontosauridae was given a formal definition in the PhyloCode by Daniel Madzia and colleagues in 2021 as "the largest clade containing Heterodontosaurus tucki, but not Iguanodon bernissartensis, Pachycephalosaurus wyomingensis, Stegosaurus stenops, and Triceratops horridus".
[2] Heterodontosaurinae is a stem-based taxon defined phylogenetically for the first time by Paul Sereno in 2012 as "the most inclusive clade containing Heterodontosaurus tucki but not Tianyulong confuciusi, Fruitadens haagarorum, Echinodon becklesii.
[4] Geranosaurus is also a heterodontosaurid, but is usually considered a nomen dubium because the type specimen is missing all its teeth, making it indistinguishable from any other genus in the family.
[25] José Bonaparte also classified the South American Pisanosaurus as a heterodontosaurid at one time,[26] but this animal is now known to be a more basal ornithischian.
[5][18] Due to supposed similarities in the morphology of the forelimbs, Robert Bakker proposed a relationship between heterodontosaurids and early sauropodomorphs like Anchisaurus, bridging the orders Saurischia and Ornithischia.
[3][4][8][28] However, others have suggested that heterodontosaurids instead share a common ancestor with Marginocephalia (ceratopsians and pachycephalosaurs),[29][30] a hypothesis that has found support in some early 21st century studies.
[34][35] In 2007, a cladistic analysis suggested that heterodontosaurids are basal to all known ornithischians except Pisanosaurus, a result that echoes some of the very earliest work on the family.
[36][37] However, a study by Bonaparte found the Pisanosauridae to be synonymous with the Heterodontosauridae and not a separate family in its own right, thereby including Pisanosaurus as a heterodontosaur.
[39] The cladogram below shows the interrelationships within Heterodontosauridae, and follows the analysis by Sereno, 2012:[40] Echinodon Fruitadens Tianyulong Lycorhinus Pegomastax Manidens Abrictosaurus Heterodontosaurus A 2020 reworking of Cerapoda by Dieudonné and colleagues recovered the animals traditionally considered 'heterodontosaurids' as a basal grouping within Pachycephalosauria, paraphyletic with respect to the traditional, dome-headed pachycephalosaurs.
This result was based on numerous skull characteristics including the dentition, and also to account for the fact that pachycephalosaur fossils are completely unknown from the Jurassic period.
The cladogram below is an abridged version of Dieudonne and colleagues' findings:[42] Chilesaurus "Heterodontosauridae" (conventional position) Thyreophora Eocursor Agilisaurus Hexinlusaurus Ornithopoda Ceratopsia Fruitadens Lycorhinus Heterodontosaurus Abrictosaurus Tianyulong Echinodon Wannanosaurus Goyocephale Prenocephale Homalocephale Stegoceras Pachycephalosaurus While originally known only from the Early Jurassic of southern Africa, heterodontosaurid remains are now known from four continents.
Early in heterodontosaurid history, the supercontinent Pangaea was still largely intact, allowing the family to achieve a near-worldwide distribution.
[10] The oldest known possible heterodontosaurid remains are a jaw fragment and isolated teeth from the Laguna Colorada Formation of Argentina, which dates back to the Late Triassic.
These remains have a derived morphology similar to Heterodontosaurus, including a caniniform with serrations on both anterior and posterior edges, as well as high-crowned maxillary teeth lacking a cingulum.
[45] The most diverse heterodontosaurid fauna comes from the Early Jurassic of southern Africa, where fossils of Heterodontosaurus, Abrictosaurus, Lycorhinus, and the dubious Geranosaurus are found.
In addition, beginning in the 1970s, a great deal of fossil material was discovered from the Late Jurassic Morrison Formation near Fruita, Colorado in the United States.
[48] The remains of Echinodon were redescribed in 2002, showing that it may represent a late-surviving heterodontosaurid from the Berriasian stage of the Early Cretaceous in southern England.
Many features of the hindlimb, including the long tibia and foot, as well as the fusion of the tibiofibiotarsus and tarsometatarsus, indicate that heterodontosaurids were adapted to run quickly on the hindlegs, so it is unlikely that Heterodontosaurus moved on all four limbs except perhaps when feeding.