lac operon
This lactose metabolism system was used by François Jacob and Jacques Monod to determine how a biological cell knows which enzyme to synthesize.
Hence the gene regulation by lac operon occurs at the transcriptional level, by preventing conversion of DNA into mRNA.
In this case, when lactose is required as a sugar source for the bacterium, the three genes of the lac operon can be expressed and their subsequent proteins translated: lacZ, lacY, and lacA.
If this weren't the case, there would be no lacY transporter protein in the cellular membrane; consequently, the lac operon would not be able to detect the presence of lactose.
Then more β-galactoside permease is synthesized allowing even more lactose to enter and the enzymes encoded by lacZ and lacA can digest it.
This dual control mechanism causes the sequential utilization of glucose and lactose in two distinct growth phases, known as diauxie.
The lac genes are organized into an operon; that is, they are oriented in the same direction immediately adjacent on the chromosome and are co-transcribed into a single polycistronic mRNA molecule.
[7] The DNA sequence of the E. coli lac operon, the lacZYA mRNA, and the lacI genes are available from GenBank (view).
If lactose is missing from the growth medium, the repressor binds very tightly to a short DNA sequence just downstream of the promoter near the beginning of lacZ called the lac operator.
Thus altered, the repressor is unable to bind to the operator, allowing RNAP to transcribe the lac genes and thereby leading to higher levels of the encoded proteins.
Transport of glucose is accompanied by its phosphorylation by EIIBGlc, draining the phosphate group from the other PTS proteins, including EIIAGlc.
One idea is that the system works through tethering; if bound repressor releases from O1 momentarily, binding to a minor operator keeps it in the vicinity, so that it may rebind quickly.
Studies have shown, that without the presence of non-specific binding, induction (or unrepression) of the Lac-operon could not occur even with saturated levels of inducer.
It had been demonstrated that, without non-specific binding, the basal level of induction is ten thousand times smaller than observed normally.
coli conserves cellular resources and energy by not making the three Lac proteins when there is no need to metabolize lactose, such as when other sugars like glucose are available.
During World War II, Monod was testing the effects of combinations of sugars as nutrient sources for E. coli and B. subtilis.
[18] Monod then focused his attention on the induction of β-galactosidase formation that occurred when lactose was the sole sugar in the culture medium.
[19] A conceptual breakthrough of Jacob and Monod[20] was to recognize the distinction between regulatory substances and sites where they act to change gene expression.
A former soldier, Jacob used the analogy of a bomber that would release its lethal cargo upon receipt of a special radio transmission or signal.
In particular, it is determined whether LacZ and LacY are made even in the absence of IPTG (due to the lactose repressor produced by the mutant gene being non-functional).
We say that the operator mutation is cis-dominant, it is dominant to wild type but affects only the copy of the operon which is immediately adjacent to it.
This explanation is misleading in an important sense, because it proceeds from a description of the experiment and then explains the results in terms of a model.
Source:[21] Explanation of diauxie depended on the characterization of additional mutations affecting the lac genes other than those explained by the classical model.
Two other genes, cya and crp, subsequently were identified that mapped far from lac, and that, when mutated, result in a decreased level of expression in the presence of IPTG and even in strains of the bacterium lacking the repressor or operator.
In a cya mutant, the absence of cAMP makes the expression of the lacZYA genes more than ten times lower than normal.
In summary: The delay between growth phases reflects the time needed to produce sufficient quantities of lactose-metabolizing enzymes.
After a delay needed to increase the level of the lactose metabolizing enzymes, the bacteria enter into a new rapid phase of cell growth.
Two puzzles of catabolite repression relate to how cAMP levels are coupled to the presence of glucose, and secondly, why the cells should even bother.
It is possible that a small difference in efficiency of transport or metabolism of glucose v. lactose makes it advantageous for cells to regulate the lac operon in this way.
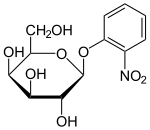
The many lac fusion techniques which include only the lacZ gene are thus suited to X-gal plates[16] or ONPG liquid broths.

1 : RNA polymerase, 2 : Repressor, 3 : Promoter, 4 : Operator, 5 : Lactose, 6 : lacZ , 7 : lacY , 8 : lacA .