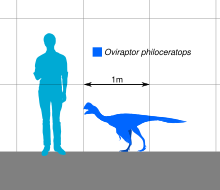
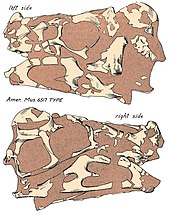
Oviraptor
The first remains were collected from the Djadokhta Formation of Mongolia in 1923 during a paleontological expedition led by Roy Chapman Andrews, and in the following year the genus and type species Oviraptor philoceratops were named by Henry Fairfield Osborn.
Despite the fact that numerous specimens have been referred to the genus, Oviraptor is only known from a single partial skeleton regarded as the holotype, as well as a nest of about fifteen eggs and several small fragments from a juvenile.
The first remains of Oviraptor were discovered on reddish sandstones of the Late Cretaceous Djadokhta Formation of Mongolia, at the Bayn Dzak locality (also known as Flaming Cliffs), during the Third Central Asiatic expedition in 1923.
This expedition was led by the North American naturalist Roy Chapman Andrews and ended in the discovery of three new-to-science theropod fossil remains—including those of Oviraptor.
These were formally described by the North American paleontologist Henry Fairfield Osborn in 1924, who in the basis of the new material, named the genera Oviraptor, Saurornithoides and Velociraptor.
[1] In 1976, the Mongolian paleontologist Rinchen Barsbold noted some inconsistencies regarding the taxonomic placement of Oviraptor and concluded that this taxon was quite distinct from ornithomimids based on anatomical traits.
[3] After Osborn's initial description of Oviraptor, the egg nest associated with the holotype was accepted to have belonged to Protoceratops,[4][5] and oviraptorids were largely considered to have been egg-eating theropods.
This new line of evidence showed that the nest associated with the holotype of Oviraptor belonged to it and the specimen was actually brooding the eggs at the time of death, not preying on them.
[11][12] A partial individual also with eggs from the Bayan Mandahu Formation of Mongolia was referred in 1996 by Dong Zhiming and Philip J. Currie, the specimen IVPP V9608.
[15][11] This conception was refuted by James M. Clark and colleagues in 2002, who noted that this tall-crested specimen has more features of the skull in common with Citipati than it does with Oviraptor—which in fact, does not preserve a crest—and it may represent a second species of the former, or, an entire new genus.
[10] However, a re-examination by Barsbold in 1997 found enough differences in this specimen to name the new genus Rinchenia, but he did not describe it with formality and this new oviraptorid remained as a nomen dubium.
[18] The North American paleontologist Mark A. Norell and colleagues in 2018 reported a new specimen of Oviraptor: AMNH 33092, which is composed of a tibia and two metatarsals of a nestling or very small juvenile.
[20][21][22] Though the holotype largely lacks the posterior region of the skeleton, it is likely that Oviraptor had two well-developed hindlimbs that ended in three functional toes with the first one being vestigial, as well as a relatively reduced tail.
[23][24] The skull of Oviraptor was deep and shortened with large fenestrae (openings) compared to other dinosaurs, and measures about 17.9 cm (179 mm) long as preserved.
He argued that the strength of the robust lower jaws and likely rhamphotheca (horny beak) was strong enough to break the shells of mollusks such as clams, which are found in the same geological formation as Oviraptor.
Barsbold also suggested that oviraptorids could have had a semiaquatic life-style based on the mollusk-based diet, the high location of nasal cavities, an augmented musculature of the tail, and the greater size of the first manual digit.
As the skull is toothless, lightly built and lacks several strong muscle insertion areas, Smith suggested that leaves may have been an important part in the diet of Oviraptor.
[31] However, in 1995, Norell and colleagues reported the fragmented remains of a lizard in the body cavity of the holotype specimen, suggesting that Oviraptor was partially carnivorous.
Based on these observations, Jansen suggested that oviraptorids were omnivorous as the sharply-developed rhamphotheca together with the prominent forelimbs would have been adapted to catch and tear small prey.
Jansen pointed out that a fully herbivorous diet in oviraptorids seems unlikely as they lacked flat and wide tomia (cutting edges of the mandibles) to chew, and were unable to move the lower jaws sideways.
The symphyseal (bone union) region at the front of the dentary may have given some ability for crushing, but as this was a relatively small area, it was probably not the main function of the jaws.
Another argument against them having been eaters of mollusks is the fact that most oviraptorids have been found in sediments that are interpreted to represent mostly arid or semi-arid environments, such as Oviraptor in the Djadokhta Formation.
[28] Thomas P. Hopp and Mark J. Orsen in 2004 analyzed the brooding behavior of extinct and extant dinosaur species, including oviraptorids, in order to evaluate the reason for the elongation and development of wing and tail feathers.
Based on the forelimb position of nesting oviraptorids, Hopp and Orsen proposed brooding as the ancestral reason behind wing and tail feather elongation, as there was a greater need to provide optimal protection for eggs and juveniles.
As most oviraptorid specimens have been found in formations of caliche-based sedimentation, Yang and colleagues suggested that the cuticle-coated eggs would have been a reproductive strategy adapted for enhancing their hatching success in such arid climates and environments.
[40] The paleoenvironment of the Djadokhta Formation is interpreted as having a semiarid climate, with sand dune and alluvial settings similar to the modern Gobi Desert.
[41][42] Though this formation is largely considered to preserved highly arid environments, several short-lived water bodies have been reported from the Ukhaa Tolgod locality, based on fluvial sedimentation.
The known remains of Oviraptor have been produced by the Bayn Dzak member, which has also yielded the dinosaurs Bainoceratops, Pinacosaurus, Protoceratops, Saurornithoides, Velociraptor,[40] and Halszkaraptor.
[44] Further dinosaur fauna from this member includes that of the Ukhaa Tolgod locality, composed of Apsaravis, Byronosaurus, Citipati, Gobipteryx, Khaan, Khol, Shuuvuia, Tsaagan,[43] and Minotaurasaurus.
[45] The pose of the holotype of Oviraptor along with the association of eggs, suggest that it was trapped over the nest during a sandstorm, and burial was relatively rapid given that the body had no opportunity to become fully disarticulated or scavenged by predators.