Wnt signaling pathway
All three pathways are activated by the binding of a Wnt-protein ligand to a Frizzled family receptor, which passes the biological signal to the Dishevelled protein inside the cell.
Its role in embryonic development was discovered when genetic mutations in Wnt pathway proteins produced abnormal fruit fly embryos.
[5] This pathway's clinical importance was demonstrated by mutations that lead to various diseases, including breast and prostate cancer, glioblastoma, type II diabetes and others.
[6][7] In recent years, researchers reported first successful use of Wnt pathway inhibitors in mouse models of disease.
[3] Since previous research by Christiane Nüsslein-Volhard and Eric Wieschaus (which won them the Nobel Prize in Physiology or Medicine in 1995) had already established the function of Wg as a segment polarity gene involved in the formation of the body axis during embryonic development, researchers determined that the mammalian int1 discovered in mice is also involved in embryonic development.
[3] Wnt comprises a diverse family of secreted lipid-modified signaling glycoproteins that are 350–400 amino acids in length.
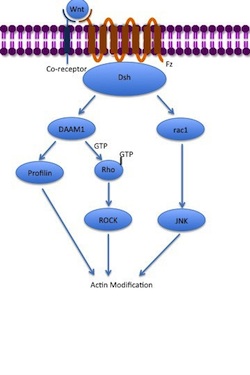
The receptor then recruits Dsh, which uses its PDZ and DIX domains to form a complex with Dishevelled-associated activator of morphogenesis 1 (DAAM1).
CaMKII induces activation of the transcription factor NFAT, which regulates cell adhesion, migration and tissue separation.
However, Wnt can also serve as a negative regulator of mTOR via activation of the tumor suppressor TSC2, which is upregulated via Dsh and GSK3 interaction.
Dsh can also interact with aPKC, Pa3, Par6 and LGl in order to control cell polarity and microtubule cytoskeleton development.
The protein porcupine mediates this process, which means that it helps regulate when the Wnt ligand is secreted by determining when it is fully formed.
[7][24] Upon secretion, the ligand can be prevented from reaching its receptor through the binding of proteins such as the stabilizers Dally and glypican 3 (GPC3), which inhibit diffusion.
[48] A cysteine-rich domain at the N-lobe of GPC3 has been identified to form a Wnt-binding hydrophobic groove including phenylalanine-41 that interacts with Wnt.
Specific antagonists include Dickkopf (Dkk), Wnt inhibitory factor 1 (WIF-1),[50][51] secreted Frizzled-related proteins (SFRP), Cerberus, Frzb, Wise, SOST, and Naked cuticle.
The synthesis of PGE2 is necessary for Wnt signaling mediated processes such as tissue regeneration and control of stem cell population in zebrafish and mouse.
[5] Intriguingly, the unstructured regions of several oversized intrinsically disordered proteins play crucial roles in regulating Wnt signaling.
In mammals, the primitive streak and other surrounding tissues produce the morphogenic compounds Wnts, BMPs, FGFs, Nodal and retinoic acid to establish the posterior region during late gastrula.
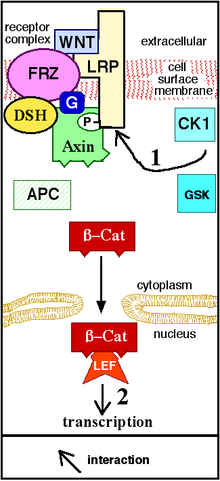
Canonical Wnt signaling β-catenin production induces the formation of this organizer via the activation of the genes twin and siamois.
[54][55] Wnt signaling is also involved in the axis formation of specific body parts and organ systems later in development.
In vertebrates, sonic hedgehog (Shh) and Wnt morphogenetic signaling gradients establish the dorsoventral axis of the central nervous system during neural tube axial patterning.
Increased β-catenin can initiate transcriptional activation of proteins such as cyclin D1 and c-myc, which control the G1 to S phase transition in the cell cycle.
These so-called Wnt-addicted cells hijack and depend on constant stimulation of the Wnt pathway to promote their uncontrolled growth, survival and migration.
LF3 strongly inhibits this binding in vitro, in cell lines and reduced tumor growth in mouse models.
[70] Cell migration during embryonic development allows for the establishment of body axes, tissue formation, limb induction and several other processes.
The fact that Wnt1 is a homolog of Wg shows that it is involved in embryonic development, which often calls for rapid cell division and migration.
[74] Its presence is revealed by elevated levels of β-catenin in the nucleus and/or cytoplasm, which can be detected with immunohistochemical staining and Western blotting.
[78] Changes in CTNNB1 expression, which is the gene that encodes β-catenin, can be measured in breast, colorectal, melanoma, prostate, lung, and other cancers.
[79] Wnt signaling is further implicated in the pathogenesis of bone metastasis from breast and prostate cancer with studies suggesting discrete on and off states.
Overexpression of Wnt5b, for instance, may increase susceptibility due to its role in adipogenesis, since obesity and type II diabetes have high comorbidity.
This leads to increased production of reactive oxygen species (ROS) known to cause DNA and cellular damage.