Wiwaxia
[4][6][7] The living animal would have measured up to 5 centimetres (2 in) when fully grown, although a range of juvenile specimens are known, the smallest being 2 millimetres (0.08 in) long.
More recently, evidence for a molluscan affinity has been accumulating, based on new details of Wiwaxia's mouthparts, scales, and growth history.
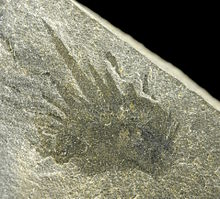
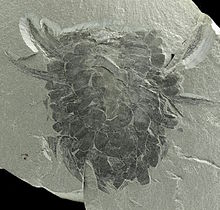
[3][6][7][10] Wiwaxia was bilaterally symmetrical; viewed from the top the body was elliptical with no distinct head or tail, and from the front or rear it was almost rectangular.
[8] The animal was covered in eight rows of small ribbed armor plates called sclerites; these lay flat against the body, overlapped so that the rear of one covered the front of the one behind, and formed five main regions—the top; the upper part of the sides; the lower part of the sides; the front; and the bottom.
Most of the sclerites were shaped like oval leaves, but the ventro-lateral ones, nearest the sea-floor, were crescent-shaped, rather like flattened bananas, and formed a single row.
Butterfield (1990) examined some sclerites under both optical and scanning electron microscopes and concluded that they were not hollow, and that the bases split and spread to form the blades, a pattern that is also seen in monocot leaves.
[16] In 1985, Simon Conway Morris agreed that there were similarities to polychaetes, but considered that Wiwaxia's sclerites were different in construction from annelids' elytra.
He was more impressed by the similarities between Wiwaxia's feeding apparatus and a molluscan radula, and assigned the animal to a new taxon Molluscata, which he proposed should also contain the molluscs and hyolithids.
[17] Nick Butterfield, then a postgraduate paleontologist at Harvard inspired by Stephen Jay Gould's lectures, agreed that the sclerites were not like elytra, which are relatively fleshy and soft.
Some modern annelids also develop on each side rows of longer bristles, which both Walcott and Butterfield considered similar to Wiwaxia's dorsal spines.
[20] Scheltema has also highlighted similarities between Wiwaxia and the larvae of certain solenogaster molluscs, which bear iterated calcareous sclerites arranged into three symmetrical lateral zones.
[21] Danish zoologist Danny Eibye-Jacobsen argued in 2004 that Wiwaxia lacks any characters that would firmly place it as a polychaete or annelid.
[16] A 2012 study redescribing the mouthparts found a number of similarities with the molluscan radula, and overthrew some of the better arguments for an annelid affinity, seemingly demonstrating that Wiwaxia was indeed a mollusc.
[8] Wiwaxia was originally described by G. F. Matthew in 1899,[23] from an isolated spine that had been found earlier in the Ogyopsis Shale, and classified as "Orthotheca corrugata".
[6] Further specimens were found by American paleontologist Charles Doolittle Walcott in 1911 as a result of one of his field trips to the nearby Burgess Shale (Miaolingian, 508 million years ago) in the Canadian Rocky Mountains; he classified it as a member of the polychaete group of annelid worms in its own genus Wiwaxia corrugata, citing similarities to the Aphroditidae and Polynoidae.
[25] Eventually in 1985 Simon Conway Morris, then a member of Whittington's team, published a detailed description and concluded that Wiwaxia was not a polychaete.
[2] What is surprising is the limited variety exhibited between species: all have a fundamentally equivalent scleritome, displaying a notable degree of morphological stasis for some 15 Ma.
[6] Younger spines of possible wiwaxiid origin have been observed from the Valongo Formation (Middle Ordovician: Dapingian-Darriwilian) of northern Portugal[1] and have been reported, if not described, from the Fezouata Biota.