Arbuscular mycorrhiza
Plants of the Rhynie chert from the Lower Devonian (400 m.yrs ago) were found to contain structures resembling vesicles and spores of present Glomus species.
The development of the AM fungi prior to root colonization, known as presymbiosis, consists of three stages: spore germination, hyphal growth, host recognition and appressorium formation.
[22][24] The branching of AM fungal hyphae grown in phosphorus media of 1 mM is significantly reduced, but the length of the germ tube and total hyphal growth were not affected.
[24] Root exudates from AMF host plants grown in a liquid medium with and without phosphorus have been shown to affect hyphal growth.
Host plant root exudates trigger and turn on AM fungal genes required for the respiration of spore carbon compounds.
It may be part of a fungal regulatory mechanism that conserves spore energy for efficient growth and the hyphal branching upon receiving signals from a potential host plant.
However, the hyphae did not further penetrate the cells and grow in toward the root cortex, which indicates that signaling between symbionts is required for further growth once appressoria are formed.
[22] Once inside the parenchyma, the fungus forms highly branched structures for nutrient exchange with the plant called arbuscules.
Trehalose and glycogen are carbon storage forms that can be rapidly synthesized and degraded and may buffer the intracellular sugar concentrations.
[37] In some cases, arbuscular mycorrhizae are poor symbionts, providing little phosphorus while taking relatively high amounts of carbon.
The hyphal NH4+ uptake also increases the nitrogen flow to the plant as the soil's inner surfaces absorb ammonium and distribute it by diffusion.
The factor was first identified by Fabienne Maillet and coworkers[40] in a groundbreaking work published in Nature, where they have extracted three hundred litre mycorrhized carrot roots and exudates from 40 million germinating spores of Rhizophagus irregularis and purified the active fraction.
Recognition of Myc factors triggers the common symbiotic signaling pathway (CSSP) that eventually leads to plant's accommodation programme to provide hostage to the arbuscular mycorrhizae.
A recent work[41] by Kevin Cope and colleagues shown that possibly other type of mycorrhizae may involve the CSSP components such as Myc-factor recognition.
Molecules similar to Nod factors were isolated from AM fungi and were shown to induce MtEnod11, lateral root formation and enhance mycorrhization.
There is also evidence to suggest that AM fungi may play an important role on mediating the plant species' specific effect on the bacterial composition of the rhizosphere.
[85] Members of the mustard family (Brassicaceae), such as cabbage, cauliflower, canola, and crambe, do not establish arbuscular mycorrhizal fungi on their roots.
[89] All the recent advances in molecular genetics clearly permit the analysis of microbial communities at much finer and functional scales and potentially with more confidence than previous methods.
Additionally, by following the traditional extraction, culture and microscopic identification methods, there is no way to determine active/functioning AM fungal populations, which are likely the most important when attempting to relate plant-AM symbiotic interactions and mechanisms to ecological or ecosystem function.
The major problem with this analysis is in field soils, which contain multiple species of AM fungi in association with a target plant at the same time (see Ecology of AM).
These AM specific primers (discussed above) can be chosen by the researcher and this decision is typically guided by the question at hand, resources available, and willingness to troubleshoot in the lab.
DNA microarray analysis is currently being used in AM fungal research to simultaneously measure the expression of many genes from target species or experimental samples.
Stable isotope probing (SIP) is a technique that can be used to determine the active metabolic function of individual taxa within a complex system of microbes.
Little has been done with this method in arbuscular mycorrhizal experiments, but if proven to work in a controlled experiment, and with further refinement of DNA/RNA fungal community analyses techniques, this may be a viable option to very specifically determine the actively growing portion of AM fungal species across growing seasons, with different plant hosts or treatments, and in the face of climate change.
Management of AM fungi is especially important for organic and low-input agriculture systems where soil phosphorus is, in general, low, although all agroecosystems can benefit by promoting arbuscular mycorrhizae establishment.
Agricultural practices such as reduced tillage, low phosphorus fertilizer usage, and perennialized cropping systems promote functional mycorrhizal symbiosis.
The disruption of the hyphal network decreases the absorptive abilities of the mycorrhizae because the surface area spanned by the hyphae is greatly reduced.
A decrease in mycorrhizal colonization due to high soil-phosphorus levels can lead to plant deficiencies in other micronutrients that have mycorrhizal-mediated uptake such as copper (Timmer & Leyden 1980).
The extraradical mycelia are able to survive the winter, providing rapid spring colonization and early season symbiosis (McGonigle and Miller 1999).
A similar decline in GRSP has also been observed in incubated soils from forested, afforested, and agricultural land[108] and grasslands treated with fungicide.




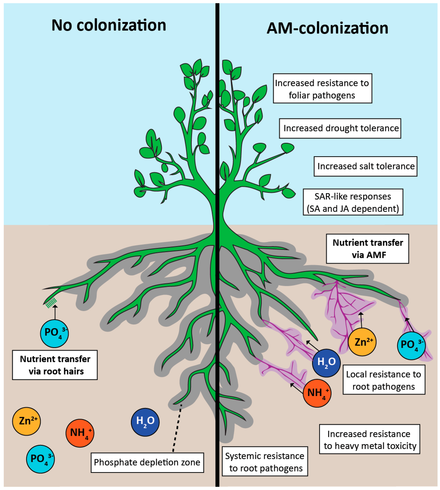


on plant performance and soil fertility
On the right: mycorrhizal helper (MH) and plant growth promoting (PGP) bacteria synergistically interacting with AMF. [ 107 ]