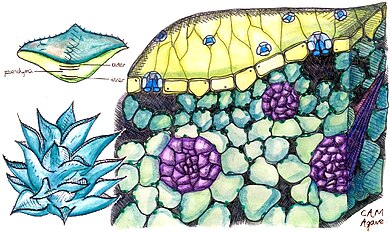
Crassulacean acid metabolism
In a plant using full CAM, the stomata in the leaves remain shut during the day to reduce evapotranspiration, but they open at night to collect carbon dioxide (CO2) and allow it to diffuse into the mesophyll cells.
The CO2 is stored as four-carbon malic acid in vacuoles at night, and then in the daytime, the malate is transported to chloroplasts where it is converted back to CO2, which is then used during photosynthesis.
Observations relating to CAM were first made by de Saussure in 1804 in his Recherches Chimiques sur la Végétation.
[3] These observations were studied further and refined by Aubert, E. in 1892 in his Recherches physiologiques sur les plantes grasses and expounded upon by Richards, H. M. 1915 in Acidity and Gas Interchange in Cacti, Carnegie Institution.
CAM is an adaptation for increased efficiency in the use of water, and so is typically found in plants growing in arid conditions.
)[6] During the night, a plant employing CAM has its stomata open, allowing CO2 to enter and be fixed as organic acids by a PEP reaction similar to the C4 pathway.
The resulting organic acids are stored in vacuoles for later use, as the Calvin cycle cannot operate without ATP and NADPH, products of light-dependent reactions that do not take place at night.
Being able to keep stomata closed during the hottest and driest part of the day reduces the loss of water through evapotranspiration, allowing such plants to grow in environments that would otherwise be far too dry.
Due to the inactivity required by the CAM mechanism, C4 carbon fixation has a greater efficiency in terms of PGA synthesis.
[11] It is now known, however, that in at least some species such as Portulaca oleracea, C4 and CAM photosynthesis are fully integrated within the same cells, and that CAM-generated metabolites are incorporated directly into the C4 cycle.
At low temperatures (frequently at night), plants using CAM open their stomata, CO2 molecules diffuse into the spongy mesophyll's intracellular spaces and then into the cytoplasm.
The latter is not possible at low temperatures, since malate is efficiently transported into the vacuole, whereas PEP-C kinase readily inverts dephosphorylation.
During the following cool night, PEP is finally exported into the cytoplasm, where it is involved in fixing carbon dioxide via malate.
Aquatic CAM plants capture carbon at night when it is abundant due to a lack of competition from other photosynthetic organisms.
This allows these trees to benefit from the elevated growth rates of C3 photosynthesis, when water is plentiful, and the drought tolerant nature of CAM, when the dry season occurs.