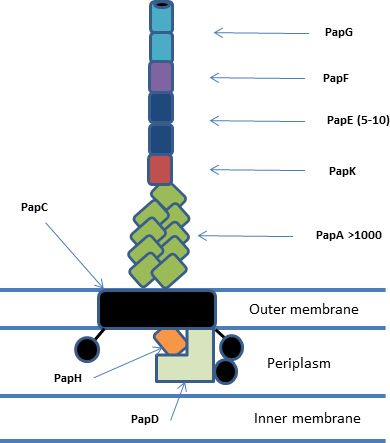
Chaperone-usher fimbriae
These fimbriae are built up out of modular pilus subunits, which are transported into the periplasm in a Sec dependent manner.
The NTD and CTDs function to bring chaperone/subunit complexes to the growing pilus and facilitate translocation across the pore.
[4] The plug domain acts to stop premature formation of a pilus and can either be located inside the pore or on the periplasmic side of the membrane.
[5] The periplasmic chaperone (PapD) has a 'boomerang' structure formed by an immunoglobulin (Ig) like fold with an essential C-terminal extension (G1).
The chaperone functions to stop aggregation and degradation of pilus subunits by binding their interfacing regions and facilitating correct protein folding before bringing them to the usher/fimbriae complex.
When the G1 is bound it binds in a parallel conformation stabilising the subunit structure but doesn't form a true Ig fold (which is anti-parallel).
FimH is the first subunit of the type I pilus (γ-1 clade) in species such as UPEC and binds to D-mannosylated residues in the bladder, allowing pathogen adhesion and colonisation.
Immune evasion is also facilitated by the tip adhesin, for example the binding of Decay accelerating factor (DAF) by gamma3 clade fimbriae inhibit effective complement activation.
By targeting pathogenicity instead of the organism (in the case of conventional antibiotics) the strong selective pressure for evolution of resistance is removed.
In the case of UPEC there have been some success in stopping pilus formation in clinical trials and the development of an effective vaccine in mice and cynomolgus monkeys based on FimCH complex.