Chloroplast
The chemical energy created is then used to make sugar and other organic molecules from carbon dioxide in a process called the Calvin cycle.
Despite this, chloroplasts can be found in extremely diverse organisms that are not directly related to each other—a consequence of many secondary and even tertiary endosymbiotic events.
The first definitive description of a chloroplast (Chlorophyllkörnen, "grain of chlorophyll") was given by Hugo von Mohl in 1837 as discrete bodies within the green plant cell.
[12][32] Chlorarachniophytes are a rare group of organisms that also contain chloroplasts derived from green algae,[29] though their story is more complicated than that of the euglenophytes.
Thiodictyon intracellulare (Chromatiaceae), a purple sulfur bacterium with a genome just half the size of their closest known relatives; and Chlorella sp.
They synthesize ordinary starch, which is stored in granules found in the periplastid space—outside the original double membrane, in the place that corresponds to the ancestral red alga's cytoplasm.
[32] However the diatom endosymbiont can't store its own food—its storage polysaccharide is found in granules in the dinophyte host's cytoplasm instead.
[12][65] The diatom endosymbiont's nucleus is present, but it probably can't be called a nucleomorph because it shows no sign of genome reduction, and might have even been expanded.
[77] While chloroplast genomes can almost always be assembled into a circular map, the physical DNA molecules inside cells take on a variety of linear and branching forms.
[85][86] The results of the microscopy experiments led to the idea that chloroplast DNA replicates using a double displacement loop (D-loop).
[23][119] In addition, in terms of function, the inner chloroplast membrane, which regulates metabolite passage and synthesizes some materials, has no counterpart in the mitochondrion.
[129] The protein-rich,[23] alkaline,[119] aqueous fluid within the inner chloroplast membrane and outside of the thylakoid space is called the stroma,[23] which corresponds to the cytosol of the original cyanobacterium.
[136] Plastoglobuli form when a bubble appears between the layers of the lipid bilayer of the thylakoid membrane, or bud from existing plastoglobuli—though they never detach and float off into the stroma.
[143][144] Thylakoids (sometimes spelled thylakoïds),[146] are small interconnected sacks which contain the membranes that the light reactions of photosynthesis take place on.
[129] Inside the photosystems embedded in chloroplast thylakoid membranes are various photosynthetic pigments, which absorb and transfer light energy.
Chl = chlorophyll[12][155][157] To fix carbon dioxide into sugar molecules in the process of photosynthesis, chloroplasts use an enzyme called RuBisCO.
This is a big problem, since O2 is produced by the initial light reactions of photosynthesis, causing issues down the line in the Calvin cycle which uses RuBisCO.
In mesophyll cells, chloroplasts are specialized for the light reactions, so they lack RuBisCO, and have normal grana and thylakoids,[140] which they use to make ATP and NADPH, as well as oxygen.
[140][159] Because the job of bundle sheath chloroplasts is to carry out the Calvin cycle and make sugar, they often contain large starch grains.
Lower levels of reactive oxygen species initiate systemic acquired resistance, triggering defense-molecule production in the rest of the plant.
[163] One of the main functions of the chloroplast is its role in photosynthesis, the process by which light is transformed into chemical energy, to subsequently produce food in the form of sugars.
Like ATP synthase, ferredoxin-NADP+ reductase, the enzyme that reduces NADP+, releases the NADPH it makes into the stroma, right where it is needed for the dark reactions.
Most of the G3P molecules are recycled back into RuBP using energy from more ATP, but one out of every six produced leaves the cycle—the end product of the dark reactions.
[166] Several mechanisms have evolved in different lineages that raise the carbon dioxide concentration relative to oxygen within the chloroplast, increasing the efficiency of photosynthesis.
[180] Other lipids are derived from the methyl-erythritol phosphate (MEP) pathway and consist of gibberelins, sterols, abscisic acid, phytol, and innumerable secondary metabolites.
[190] Studies of Vallisneria gigantea, an aquatic flowering plant, have shown that chloroplasts can get moving within five minutes of light exposure, though they don't initially show any net directionality.
An etioplast is a plastid that lacks chlorophyll, and has inner membrane invaginations that form a lattice of tubes in their stroma, called a prolamellar body.
[188][196] Late into the constriction phase, dynamin proteins assemble around the outer plastid-dividing ring,[196] helping provide force to squeeze the chloroplast.
There are many other documented mechanisms that prevent paternal inheritance in these flowering plants, such as different rates of chloroplast replication within the embryo.
This makes plastid transformation a valuable tool for the creation and cultivation of genetically modified plants that are biologically contained, thus posing significantly lower environmental risks.


a It is now established that Chromalveolata is paraphyletic to Rhizaria . [ 32 ]








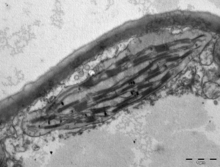



(Top) 10-nm-thick STEM tomographic slice of a lettuce chloroplast. Grana stacks are interconnected by unstacked stromal thylakoids, called "stroma lamellae". Round inclusions associated with the thylakoids are plastoglobules. Scalebar=200 nm. See. [ 145 ]
(Bottom) Large-scale 3D model generated from segmentation of tomographic reconstructions by STEM. grana=yellow; stroma lamellae=green; plastoglobules=purple; chloroplast envelope=blue. See. [ 145 ]



