Coincidence detection in neurobiology
Coincidence detectors influence neuronal information processing by reducing temporal jitter and spontaneous activity, allowing the creation of variable associations between separate neural events in memory.
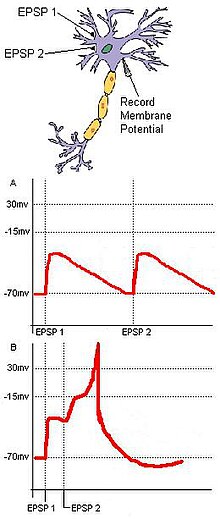
Hence, the function of coincidence detection is to reduce the jitter caused by spontaneous neuronal activity, and while random sub-threshold stimulations from cells may not often fire coincidentally, coincident synaptic inputs derived from a unitary external stimulus ensure that a target neuron will fire as a result of the stimulus.
If sufficient signals are received within a short period of time (i.e. before the overall voltage decays to background), the voltage of the segment will rise above a threshold, giving rise to a non-linear dendritic spike, which travels, effectively undiminished, all the way to the cell body, and which causes it to become partially depolarised.
Coincidence detection has been shown to be a major factor in sound localization along the azimuth plane in several organisms.
In 1948, Lloyd A. Jeffress proposed that some organisms may have a collection of neurons that receive auditory input from each ear.
Jeffress claimed that the neurons that the delay lines link act as coincidence detectors by firing maximally when receiving simultaneous inputs from both ears.
Jeffress' model proposes that two signals even from an asynchronous arrival of sound in the cochlea of each ear will converge synchronously on a coincidence detector in the auditory cortex based on the magnitude of the ITD (Fig.
A large elevation of calcium levels activate protein kinases that ultimately increase the number of postsynaptic AMPA receptors.
Besides the NMDA-receptor based processes, further cellular mechanisms allow of the association between two different input signals converging on the same neuron, in a defined timeframe.
Upon a simultaneous increase in the intracellular concentrations of cAMP and Ca2+, a transcriptional coactivator called TORC1 (CRTC1) becomes activated, that converts the temporal coincidence of the two second messengers into long term changes such as LTP.
[7] This cellular mechanism, through calcium-dependent adenylate cyclase activation, might also account for the detection of the repetitive stimulation of a given synapse.
The Ca2+, IP3, and DAG work together in a signal transduction pathway to internalize AMPA receptors and decrease the sensitivity of the postsynaptic cell to glutamate.