Drosophila embryogenesis
The study of its embryogenesis unlocked the century-long puzzle of how development was controlled, creating the field of evolutionary developmental biology.
Drosophila display a holometabolous method of development, meaning that they have three distinct stages of their post-embryonic life cycle, each with a radically different body plan: larva, pupa and finally, adult.
About 24 hours after fertilization, an egg hatches into a larva, which undergoes three molts taking about 5.5 to 6 days, after which it is called a pupa.
The entire growth process from egg to adult fly takes an estimated 10 to 12 days to complete at 25 °C.
Embryogenesis in Drosophila is unique among model organisms in that cleavage occurs in a multinucleate syncytium (strictly a coenocyte).
After thirteen mitotic divisions and about 4 hours after fertilization, an estimated 6,000 nuclei accumulate in the unseparated cytoplasm of the oocyte before they migrate to the surface and are encompassed by plasma membranes to form cells surrounding the yolk sac producing a cellular blastoderm.
The posterior region (including the hindgut) expands and extends towards the anterior pole along the dorsal side of the embryo.
Germ band retraction returns the hindgut to the dorsal side of the posterior pole and coincides with overt segmentation.
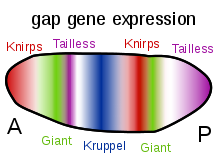
Bicoid and Hunchback are the maternal effect genes that are most important for patterning of anterior parts (head and thorax) of the Drosophila embryo.
Nanos and Caudal are maternal effect genes that are important in the formation of more posterior abdominal segments of the Drosophila embryo.
[4] Cytoskeletal elements such as microtubules are polarized within the oocyte and can be used to allow the localization of mRNA molecules to specific parts of the cell.
Maternally synthesized bicoid mRNAs attach to microtubules and are concentrated at the anterior ends of forming Drosophila eggs.
hunchback and caudal mRNAs lack special location control systems and are fairly evenly spread throughout the entire interior of the egg cells.
The caudal protein then activates later to turn genes on to form the posterior structures during the segmentation phase.
In order for development to continue, Hunchback is needed in an area that is declining in amount from anterior to posterior.
Their expression patterns in the early embryo are determined by the maternal effect gene products and shown in the diagrams on the right side of this page.
The Naked cuticle protein is induced by Wingless to limit the number of rows of cells that express Engrailed.
The short-range, reciprocal signaling by Hedgehog and Wingless, held in check by the Patched and Naked proteins, stabilizes the boundary between each segment.
In 1995, the Nobel Prize for Physiology or Medicine was awarded for studies concerning the genetic control of early embryonic development to Christiane Nüsslein-Volhard, Edward B. Lewis and Eric Wieschaus.
Their research on genetic screening for embryo patterning mutants revealed the role played in early embryologic development by homeobox genes like bicoid.
Formation of the dorsal-ventral axis is dependent on the ventral nuclear concentration of a maternally synthesized transcription factor called Dorsal.
The nucleus expresses a protein called Gurken which is secreted locally and thus only activates follicle cells in the dorsal region by interacting with the Torpedo receptor.
Toll signaling results in the degradation of Cactus which allows Dorsal to enter the nuclei on the ventral side of the blastoderm.
In the lateral regions of the embryo, low nuclear concentrations of Dorsal lead to the expression of rhomboid which identifies future neuroectoderm.
The ventral activity of the TGF-β family signaling protein Dpp is maintained by the expression of the secreted Dpp-antagonist Sog (short gastrulation) in the neuroectoderm.