Decapentaplegic
[1] It is known to be necessary for the correct patterning and development of the early Drosophila embryo and the fifteen imaginal discs, which are tissues that will become limbs and other organs and structures in the adult fly.
Flies with mutations in decapentaplegic fail to form these structures correctly, hence the name (decapenta-, fifteen; -plegic, paralysis).
Studies of Dpp in Drosophila have led to greater understanding of the function and importance of their homologs in vertebrates like humans.
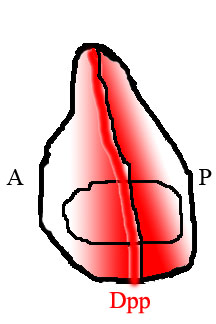
During embryonic development, Dpp is uniformly expressed at the dorsal side of the embryo, establishing a sharp concentration gradient.
The initial discovery of MAD in Drosophila paved the way for later experiments that identified the responder to TGF-β signaling in vertebrates, called SMADs.
[13] Activated MAD is able to bind to DNA and act as a transcription factor to affect the expression of different genes in response to Dpp signaling.
The result is that Dpp is only produced in a narrow stripe of cells immediately adjacent to but not within the posterior half of the tissue.
[15] Dpp produced at this anterior/posterior border then diffuses out to the edges of the tissue, forming a spatial concentration gradient.
By reading their position along the gradient of Dpp, cells in the wing are able to determine their location relative to the anterior/posterior border, and they behave and develop accordingly.
[17] Dpp has also been proposed as a regulator of tissue growth and size, a classic problem in development.
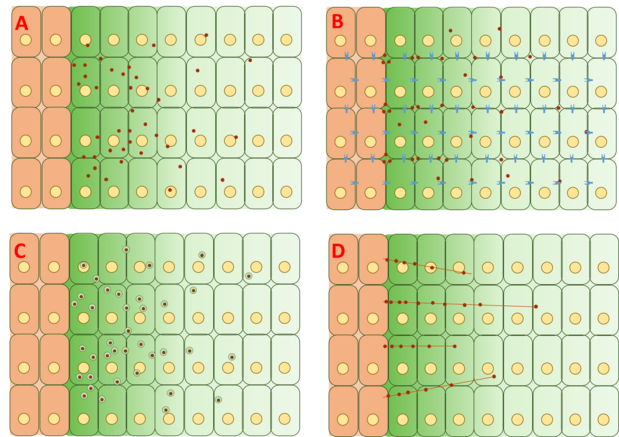
However, the mechanism by which the Dpp gradient is formed is still controversial, and no complete explanation has been proposed or proven.
FRAP assays have argued against this model by noting that diffusion of GFP-Dpp does not match that expected of a similarly sized molecule.
The restricted diffusion model includes the effects of cell packing geometry and interactions with the extracellular matrix via binding events with receptors such as Tkv and the heparin sulfate proteoglycans dally and dally-like.
Dpp fails to move across cells with mutated dally and dally-like, two heparin sulfate proteoglycans (HSPGs) commonly found in the extracellular matrix.
Dpp is also found in molluscs, where it plays a key role in shell formation by controlling the shape of the conch.