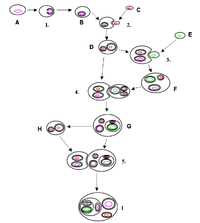
Endosymbiont
Examples are nitrogen-fixing bacteria (called rhizobia), which live in the root nodules of legumes, single-cell algae inside reef-building corals, and bacterial endosymbionts that provide essential nutrients to insects.
Roughly 2.2 billion years ago an archaeon absorbed a bacterium through phagocytosis, that eventually became the mitochondria that provide energy to almost all living eukaryotic cells.
Approximately 1 billion years ago, some of those cells absorbed cyanobacteria that eventually became chloroplasts, organelles that produce energy from sunlight.
[4] Approximately 100 million years ago, a lineage of amoeba in the genus Paulinella independently engulfed a cyanobacteria that evolved to be functionally synonymous with traditional chloroplasts, called chromatophores.
[5] Some 100 million years ago, UCYN-A, a nitrogen-fixing bacterium, became an endosymbiont of the marine alga Braarudosphaera bigelowii, eventually evolving into a nitroplast, which fixes nitrogen.
[6] Similarly, diatoms in the family Rhopalodiaceae have cyanobacterial endosymbionts, called spheroid bodies or diazoplasts, which have been proposed to be in the early stages of organelle evolution.
[10][11] Some human parasites, e.g. Wuchereria bancrofti and Mansonella perstans, thrive in their intermediate insect hosts because of an obligate endosymbiosis with Wolbachia spp.
The original Hodgkinia genome split into three much simpler endosymbionts, each encoding only a few genes—an instance of punctuated equilibrium producing distinct lineages.
Secondary endosymbionts more recently associated with their hosts, may be horizontally transferred, live in the hemolymph of the insects (not specialized bacteriocytes, see below), and are not obligate.
[34] Primary endosymbionts are thought to help the host either by providing essential nutrients or by metabolizing insect waste products into safer forms.
For example, the putative primary role of Buchnera is to synthesize essential amino acids that the aphid cannot acquire from its diet of plant sap.
Bacteria benefit from the reduced exposure to predators and competition from other bacterial species, the ample supply of nutrients and relative environmental stability inside the host.
Primary endosymbionts of insects have among the smallest of known bacterial genomes and have lost many genes commonly found in closely related bacteria.
Research in which a parallel phylogeny of bacteria and insects was inferred supports the assumption hat primary endosymbionts are transferred only vertically.
For example, aphids are crop pests and the tsetse fly carries the organism Trypanosoma brucei that causes African sleeping sickness.
[38] The pea aphid (Acyrthosiphon pisum) contains at least three secondary endosymbionts, Hamiltonella defensa, Regiella insecticola, and Serratia symbiotica.
In Drosophila neotestacea, S. poulsonii has spread across North America owing to its ability to defend its fly host against nematode parasites.
[44] Sodalis glossinidius is a secondary endosymbiont of tsetse flies that lives inter- and intracellularly in various host tissues, including the midgut and hemolymph.
These marine worms are nutritionally dependent on their symbiotic chemoautotrophic bacteria lacking any digestive or excretory system (no gut, mouth, or nephridia).
[59] Additionally, both host and symbiont cell growth were much greater than free-living Richelia intracellularis or symbiont-free Hemiaulus spp.
[54] There are some asymbiotic (occurs without an endosymbiont) Rhizosolenia, however there appears to be mechanisms limiting growth of these organisms in low nutrient conditions.
In 1966, biologist Kwang W. Jeon found that a lab strain of Amoeba proteus had been infected by bacteria that lived inside the cytoplasmic vacuoles.
[75][76][77] The spotted salamander (Ambystoma maculatum) lives in a relationship with the algae Oophila amblystomatis, which grows in its egg cases.
Endophytes aid in processes such as growth and development, nutrient uptake, and defense against biotic and abiotic stresses like drought, salinity, heat, and herbivores.
Typically, plant endosymbiosis studies focus on a single category or species to better understand their individual biological processes and functions.
[87] Plant root exudates contain diverse secondary metabolites, especially flavonoids and strigolactones that act as chemical signals and attracts the AMF.
[90] AMF generally promote plant health and growth and alleviate abiotic stresses such as salinity, drought, heat, poor nutrition, and metal toxicity.
[93] Fungal endophytes are taxonomically diverse and are divided into categories based on mode of transmission, biodiversity, in planta colonization and host plant type.
[96] Aureobasidium and preussia species of endophytic fungi isolated from Boswellia sacra produce indole acetic acid hormone to promote plant health and development.
Nevertheless, archaea are a substantial constituent of plant-associated ecosystems in the aboveground and belowground phytobiome, and play a role in host plant's health, growth and survival amid biotic and abiotic stresses.