Erlikosaurus
It represents the second therizinosaur taxon from this formation (alongside Enigmosaurus and Segnosaurus) with the most complete skull among members of this peculiar family of dinosaurs.
Like other therizinosaurids, Erlikosaurus had a large gut for food processing, strong arms ending in elongated claws, and a backwards directed pelvis.
The generic name, Erlikosaurus, was taken from that of the demon king Erlik, from Turko-Mongolian mythology and the Greek σαῦρος (sauros, meaning lizard).
At the time of its discovery it was the only known therizinosaur (then called segnosaurs[6]) for which a complete skull had been discovered, and this helped shed light on a puzzling and poorly known group of dinosaurs.
[8] However, since the holotype hip of Enigmosaurus did not closely resemble that of the specimen in Segnosaurus as would be expected for the Segnosaurus-like remains of Erlikosaurus, and there is a considerable size difference, paleontologist Rinchen Barsbold disputed the alleged synonymy.
A fine, vertical lamina of bone is connected rostrally to the medial margin of the premaxilla, indicating that when the animal was alive, a cartilaginous internasal septum was present.
[18] The preserved rhamphotheca in specimens of Gallimimus and Ornithomimus evidences that the keratin sheath covered the premaxilla and overlapped it on the lower side by a few millimeters.
[18] Body remains of Erlikosaurus are very sparse compared to the cranial elements, consisting of a humerus, a right foot and some cervical vertebrae.
The metatarsal I is the shortest in comparison, it measures 7 cm (70 mm) long and expands the laterally extended proximal articular surface of the metatarsus.
[3] With the discovery and description of the feathered Beipiaosaurus, therizinosaurs were utterly recognized as theropods, and started to be reconstructed in an accurate, bipedal posture.
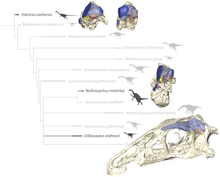
Erlikosaurus occupied a very derived position in a clade with the two Nothronychus species:[11] Suzhousaurus Neimongosaurus Therizinosaurus Erliansaurus Nanchao embryos Nanshiungosaurus Segnosaurus
Erlikosaurus Nothronychus graffami Nothronychus mckinleyi Erlikosaurus is poorly known from postcranial material, but the holotype skull became the focus of study in Computed Tomography (CT) scans that were published back in 2012 by the paleontologist Stephan Lautenschlager and Dr Emily Rayfield of Bristol University School of Earth Sciences, Professor Lindsay Zanno of the North Carolina Museum of Natural History and North Carolina State University, and Lawrence Witmer, Chang Professor of Paleontology at the Ohio University Heritage College of Osteopathic Medicine.
Analysis of the brain cavity revealed that Erlikosaurus, and quite likely most other therizinosaurids, had well developed senses of smell, hearing, and balance, traits better associated with carnivorous theropods.
These senses were also well-developed in earlier coelurosaurs and other theropods, indicating that therizinosaurs may have inherited many of these traits from their carnivorous ancestors and used them for their different and specialized dietary purposes.
The scores of Erlikosaurus were higher than most dromaeosaurids despite the herbivorous life-style in this taxon, and may reflect a transition to complex sociality and/or reduced visual capacities.
Hughes and Finarelli pointed out that as dinosaurian lineages became larger, the size of the olfactory bulb increased, which may suggest olfaction as the main sensory modality in large-bodied non-avian dinosaurs.
[28] In 2013, Lautenschlager performed digital reconstructions for the cranial musculature in Erlikosaurus and found a relatively weak bite force compared to other theropods.
Lautenschlager found the lowest force at the snout tip with 43–65 N, and the highest at the last maxillar tooth region, with 90–134 N. Factors like the presence of a large gut to process vegetation and the lack of damage patterns on the teeth suggest that Erlikosaurus used only the tip of the snout and the premaxillary region to reach for soft foliage or fruits, and the lesser bite force for Erlikosaurus better served in leaf-stripping and plant-cropping feeding mechanism, rather than active mastication.
In this study, Lautenschlager also suggested that Erlikosaurus may have been able to process mainly thin branches and plant matter based on Stegosaurus.
[30] The well preserved jaws also allowed a study by the University of Bristol to determine how its feeding style and dietary preferences were linked to how wide they could open the mouth.
The resulting bite force for Erlikosaurus was between 50 and 121 N, with a skull characterised by high susceptibility to stress and deformation that indicates a feeding behaviour specialized in the active use of the beak.
The results further support that Erlikosaurus relied on postcranial musculature to compensate the low bite force and to relieve stresses on cranial structure.
[33][3][34] The examination of the magnetostratigraphy of the formation seems to confirm that the entire Bayan Shireh lies within the Cretaceous Long Normal, which lasted only until the end of the Santonian stage.
[34][35] Fluvial, lacustrine and caliche-based sedimentation indicates a lesser semi-arid climate, with the presence of wet environments composed of large meanders and lakes.
[39] Other herbivorous dinosaurs are represented by the ankylosaurs Talarurus and Tsagantegia,[40] small marginocephalians Amtocephale and Graciliceratops,[41][42] the hadrosauroid Gobihadros,[10] and the sauropod Erketu.
In 2016, Zanno and colleagues re-examined the lower jaws and dentition of Segnosaurus making direct comparisons with those of Erlikosaurus in the process.
The respective indistinct and specialized dentition of Erlikosaurus and Segnosaurus indicates that these two therizinosaurids were separated by niche differentiation in food acquisition, processing, or resources.
In contrast, Erlikosaurus and Segnosaurus were aided by the down-turned tip of the lower jaws and symphyseal (bone union) regions, and probably also by stress and strain-mitigating beaks.
While Segnosaurus was adapted to use its specialized dentition to procure or process food, Erlikosaurus mostly relied on its beak and neck musculature for cropping while foraging.
Segnosaurus was found to be in the former mode, whereas Erlikosaurus was more likely to fall in the second group, further supporting that these two therizinosaurids were separated by a well-defined niche differentiation.