Genetic drift
In the middle of the 20th century, vigorous debates occurred over the relative importance of natural selection versus neutral processes, including genetic drift.
In 1968, population geneticist Motoo Kimura rekindled the debate with his neutral theory of molecular evolution, which claims that most instances where a genetic change spreads across a population (although not necessarily changes in phenotypes) are caused by genetic drift acting on neutral mutations.
Among the four that survive, 16 possible combinations for the A and B alleles exist: (A-A-A-A), (B-A-A-A), (A-B-A-A), (B-B-A-A), (A-A-B-A), (B-A-B-A), (A-B-B-A), (B-B-B-A), (A-A-A-B), (B-A-A-B), (A-B-A-B), (B-B-A-B), (A-A-B-B), (B-A-B-B), (A-B-B-B), (B-B-B-B).
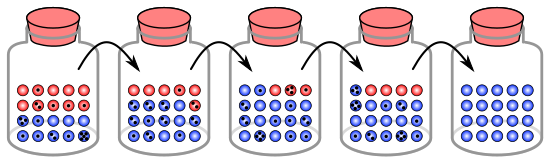
In the latter case, genetic drift has occurred because the population's allele frequencies have changed due to random sampling.
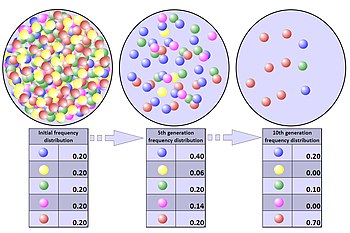
Mathematical models of genetic drift can be designed using either branching processes or a diffusion equation describing changes in allele frequency in an idealised population.
On the other hand, computer simulations are usually easier to perform using the Wright–Fisher model, because fewer time steps need to be calculated.
[2] The Hardy–Weinberg principle states that within sufficiently large populations, the allele frequencies remain constant from one generation to the next unless the equilibrium is disturbed by migration, genetic mutations, or selection.
In both cases, the time to fixation is dominated by mutation via the term 1/m, and is less affected by the effective population size.
Non-adaptive evolution resulting from the product of mutation and genetic drift is therefore considered to be a consequential mechanism of evolutionary change primarily within small, isolated populations.
[17] Genetic linkage to other genes that are under selection can reduce the effective population size experienced by a neutral allele.
Not until the allele frequency for the advantageous mutation reaches a certain threshold will genetic drift have no effect.
An interesting example of a bottleneck causing unusual genetic distribution is the relatively high proportion of individuals with total rod cell color blindness (achromatopsia) on Pingelap atoll in Micronesia.
[2] For recessive harmful mutations, this selection can be enhanced as a consequence of the bottleneck, due to genetic purging.
In addition, a sustained reduction in population size increases the likelihood of further allele fluctuations from drift in generations to come.
A population's genetic variation can be greatly reduced by a bottleneck, and even beneficial adaptations may be permanently eliminated.
[38] The loss of variation leaves the surviving population vulnerable to any new selection pressures such as disease, climatic change or shift in the available food source, because adapting in response to environmental changes requires sufficient genetic variation in the population for natural selection to take place.
The declines in population resulted from hunting and habitat destruction, but a consequence has been a loss of most of the species' genetic diversity.
Their resulting decline in genetic variation can be deduced by comparing it to that of the southern elephant seal, which were not so aggressively hunted.
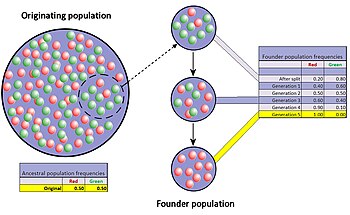
The random sample of alleles in the just formed new colony is expected to grossly misrepresent the original population in at least some respects.
When a newly formed colony is small, its founders can strongly affect the population's genetic make-up far into the future.
[30][45] The difference in gene frequencies between the original population and colony may also trigger the two groups to diverge significantly over the course of many generations.
This potential for relatively rapid changes in the colony's gene frequency led most scientists to consider the founder effect (and by extension, genetic drift) a significant driving force in the evolution of new species.
Sewall Wright was the first to attach this significance to random drift and small, newly isolated populations with his shifting balance theory of speciation.
[46] Following after Wright, Ernst Mayr created many persuasive models to show that the decline in genetic variation and small population size following the founder effect were critically important for new species to develop.
[47] However, there is much less support for this view today since the hypothesis has been tested repeatedly through experimental research and the results have been equivocal at best.
[48] The role of random chance in evolution was first outlined by Arend L. Hagedoorn and Anna Cornelia Hagedoorn-Vorstheuvel La Brand in 1921.
[57] In the early days of the modern evolutionary synthesis, scientists were beginning to blend the new science of population genetics with Charles Darwin's theory of natural selection.
To Fisher, viewing the process of evolution as a long, steady, adaptive progression was the only way to explain the ever-increasing complexity from simpler forms.
But the debates have continued between the "gradualists" and those who lean more toward the Wright model of evolution where selection and drift together play an important role.
[5][6] The role of genetic drift by means of sampling error in evolution has been criticized by John H. Gillespie[61] and William B. Provine, who argue that selection on linked sites is a more important stochastic force.