Potyvirus
Like begomoviruses, members of this genus may cause significant losses in agricultural, pastoral, horticultural, and ornamental crops.
[citation needed] The genus contains 190 species and potyviruses account for about thirty percent of all currently known plant viruses.
The typical base compositions of some of the most common, non-recombinant strains of the type species, PVY, range between ~23.4-23.8 % G; ~31-31.6 % A; ~18.2-18.8 % C; and ~26.5-26.8 % U.
The genome encodes a single open reading frame (ORF) expressed as a 350 kDa polyprotein precursor.
The P3 cistron also contains an overlapping reading frame called "Pretty interesting Potyviridae ORF" (PIPO).
[4] PIPO codes for an alternative C-terminus to the P3 protein, which is generated into a subset of transcripts by a +2 frameshift caused by a ribosome slippage mechanism at a conserved GA6 repeat sequence.
[7] P1 (~33 kilodaltons (kDa) in molecular weight) is a serine protease which facilitates its own cleavage from the polyprotein at the P1-HC-Pro junction.
[11] Though the exact mechanism is unknown, HC-Pro has been proposed to attach to host aphid mouth parts through its N-terminal zinc finger-like domain and anchor virions through its interactions with the capsid protein.
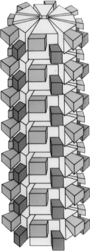
[15] Its most unusual property is its ability to form large and highly symmetrical conical and cylindrical inclusions with a central hollow cylinder from which laminate sheets radiate outward and fold in on themselves in a pattern often described as "pinwheels".
[22] Like NIa, NIb forms inclusions in the host nucleus where it is transported due to its two nuclear localisation sequences.
[24] It interacts with various ER exit site proteins to produce vesicular and tubular extensions which eventually mature into replication vesicles.
In addition to encapsidating the virion, CP core domain is required for intercellular movement and contributes to seed transmission.
[35] Vegetative transmission by infected tubers or grafting material are of particular concern for certain agricultural crops, such as potato and fruit trees, respectively.
Transmission can also occur by physical contact with infected plants or with contaminated tools, clothes, or even water.
Potyviral RNA mimicks host mRNA: the 5' VPg protein shares functional similarities with the 5' cap and the 3' end is polyadenylated.
Replication vesicle membranes are derived from a variety of host organelles and the sources differ between potyvirus species.
The exact replication mechanism is not known but it involves a negative sense RNA intermediate and requires both viral and host proteins.
Viral proteins detected in replication complexes include HC-Pro, P3, 6K1, 6K2, CI, VPg, NIa-Pro, and NIb.
[39] Host factors present in replication vesicles include eIF4A and several heat shock proteins.
[citation needed] Historically, potyvirus diagnostics relied on the detection of various proteinaceous inclusions in infected plant cells.
Gibbs and Ohshima 2010 produced a more extensive molecular phylogeny with the same four, but also several new groups: the BtMV, ChVMV, DaMV, OYDV, PRSV, TEV, and TuMV.