Hedgehog signaling pathway
Hh is one of Drosophila's segment polarity gene products, involved in establishing the basis of the fly body plan.
In knockout mice lacking components of the pathway, the brain, skeleton, musculature, gastrointestinal tract and lungs fail to develop correctly.
[1] Drugs that specifically target Hedgehog signaling to fight this disease are being actively developed by a number of pharmaceutical companies.
In the 1970s, a fundamental problem in developmental biology was to understand how a relatively simple egg can give rise to a complex segmented body plan.
In the late 1970s Christiane Nüsslein-Volhard and Eric Wieschaus isolated mutations in genes that control development of the segmented anterior-posterior body axis of the fly;[4] their "saturation mutagenesis" technique resulted in the discovery of a group of genes involved in the development of body segmentation, helping to found the field of evolutionary developmental biology.
The fly hh gene was independently cloned in 1992 by the labs of Jym Mohler, Philip Beachy, Thomas B. Kornberg and Saigo Kaoru.
Insect cells express a full size zinc-finger transcription factor Cubitus interruptus (Ci), which forms a complex with the kinesin-like protein Costal-2 (Cos2) and is localized in the cytoplasm bound to cellular microtubules (Figure 2).
For other Hh-regulated genes, expression requires not only the loss of CiR but also the positive action of uncleaved Ci to act as a transcriptional activator.
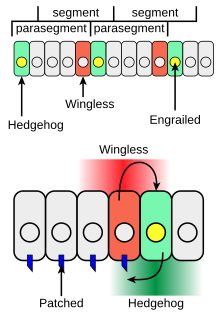
During the formation of body segments in the developing Drosophila embryo, stripes of cells that synthesize the transcription factor engrailed can also express the cell-to-cell signaling protein Hedgehog (green in Figure 4).
During the cleavage, a cholesterol molecule is added to the carboxyl end of the N-terminal domain,[18] which is involved in trafficking, secretion and receptor interaction of the ligand.
Secretion and consequent paracrine hedgehog signaling require the participation of Dispatched (DISP) protein (Process "2" on Figure 5).
[22] Upon binding of a Hh protein or a mutation in the SSD of PTCH, the pump is turned off allowing oxysterols to accumulate around SMO.
[27] Consequently, in vivo the passage of hedgehog over a receptive field that expresses the receptor leads to attenuation of the signal, an effect called ligand-dependent antagonism (LDA).
The classic experiments[29] of Saunders and Gasseling in 1968[30] on the development of the chick limb bud formed the basis of the morphogen concept.
The current model, proposed by Harfe et al.,[31] states that both the concentration and the time of exposure to SHH determines which digit the tissue will develop into in the mouse embryo (figure 6).
In these cells SHH signals in an autocrine fashion and these digits develop correctly in the absence of DISP, which is required for extracellular diffusion of the ligand.
[36] Disruption of hedgehog signaling during embryonic development, through either deleterious mutation or consumption of teratogens by the gestating mother, can lead to severe developmental abnormalities.
Holoprosencephaly, the failure of the embryonic prosencephalon to divide to form cerebral hemispheres, occurs with a frequency of about 1 in 8,000 live births and about 1 in 200 spontaneous abortions in humans and is commonly linked to mutations in genes involved in the hedgehog pathway, including SHH and PTCH.
[38] Activation of the hedgehog pathway has been implicated in the development of cancers in various organs, including brain, lung, mammary gland, prostate and skin.
Basal cell carcinoma, the most common form of cancerous malignancy, has the closest association with hedgehog signaling.
Cancer researchers hope that specific inhibitors of hedgehog signaling will provide an efficient therapy for a wide range of malignancies.
Nevertheless, it is clear that the aberrant activation of hedgehog signaling leads to the growth, proliferation, and invasion of tumor cells.
[46] Itraconazole (ITZ) inhibits SMO in the presence of mutations conferring resistance to vismodegib and other cyclopamine-competitive antagonists, like IPI-926 and Novartis' LDE-225.
[50] Arsenic trioxide (Trisenox) has also been shown to inhibit hedgehog signaling by interfering with Gli function and transcription.
Piperonyl butoxide (PBO) is a semisynthetic pesticide additive developed in the 1940s, which can be found in thousands of household and agricultural products.
[55] Despite its widespread use, the ability of PBO to inhibit hedgehog signaling and act as a potent developmental teratogen was not recognized until recently.
[56][57] Activation of the Hedgehog pathway leads to an increase in Snail protein expression and a decrease in E-cadherin and tight junctions.
Vertebrates, on the other hand, have several Hedgehog ligands that fall within three subgroups – Desert, Indian and Sonic, each represented by a single mammalian gene.
[75] The Hog domain is present in many eukaryotic branches, i.e. red algae, mosses, dinoflagellates, jakobids, and other single cell eurkaryotes.
[76][77] These N-terminal domains associated with Hog in C. elegans were subsequently classified, initially Warthog (WRT) and Groundhog (GRD), followed by Ground-like (GRL) and Quahog (QUA).