Pachycephalosauria
This can make taxonomic identification a difficult task, as the classification of genera and species within Pachycephalosauria relies almost entirely on cranial characteristics.
For instance, Majungatholus, once thought to be a pachycephalosaur, is now recognized as a specimen of the abelisaurid theropod Majungasaurus, and Yaverlandia, another dinosaur initially described as a pachycephalosaurid, has also since been reclassified as a coelurosaur (Naish in Sullivan (2006)).
[11] The same conclusion had previously been reached by George Olshevsky in 1991, who classified heterodontosaurids as basal pachycephalosaurs on the basis of perceived cranial kinesis, the presence of fanglike premaxillary teeth, and the prominent diastema present in many genera.
Sullivan considered attempts by Maryańska and Osmólska to restrict the definition of Pachycephalosauria redundant with their Pachycephalosauridae, since they were diagnosed by the same anatomical characters.
Sullivan also pointed out that the original diagnosis of Pachycephalosauridae centered around "flat to dome-like" skulls, so the flat-headed forms should be included in the family.
[16] In a paper published in 2003, Thomas E. Williamson and Thomas D. Carr discovered a clade of the Pachycephalosauridae that was a sister taxa to the genus Stegoceras, made up of "all other dome-headed pachycephalosaurians; this was referred to as Pachycephalosaurinae[17] Phylogenetic analyses by many authors have found Pachycephalosauria to be a group with Stegoceras as one of the earliest fully-domed members, with flat-headed and potentially juvenile taxa like Homalocephale and Goyocephale either just outside or just within the clade of it and more derived pachycephalosaurs.
These studies began with the phylogenetic work of Paul Sereno, which has been modified in many iterations to include newer taxa and additional characters.
[21] Wannanosaurus Hanssuesia Colepiocephale Stegoceras validum Stegoceras novomexicanum Goyocephale Homalocephale Tylocephale Foraminacephale Amtocephale Acrotholus Prenocephale Alaskacephale Stygimoloch Pachycephalosaurus S. goodwini S. lyonsi S. triregnum S. buchholtzae S. edmontonensis Below is a cladogram published by Dieudonné and colleagues (2020)[22] which controversially found heterodontosauridae to be paraphyletic with respect to pachycephalosauria.
This analysis was proposed as a hypothesis for the complete lack of Jurassic and Early Cretaceous pachycephalosaur fossils, even though they should have existed if the modern understanding of ornithischian phylogeny is correct.
[1] Fruitadens Lycorhinus Heterodontosaurus Abrictosaurus Tianyulong Echinodon Wannanosaurus Goyocephale Prenocephale Homalocephale Stegoceras Pachycephalosaurus The small size of most pachycephalosaur species and lack of skeletal adaptation indicates that they were not climbers and primarily ate food close to the ground.
Mallon et al. (2013) examined herbivore coexistence on the island continent of Laramidia, during the Late Cretaceous and concluded that pachycephalosaurids were generally restricted to feeding on vegetation at, or below, the height of 1 meter.
Wear patterns on the teeth vary by species, indicating a range of food preferences which could include seeds, stems, leaves, fruits, and possibly insects.
A very wide rib cage and large gut cavity extending all the way to the base of the tail suggests the use of fermentation to digest food.
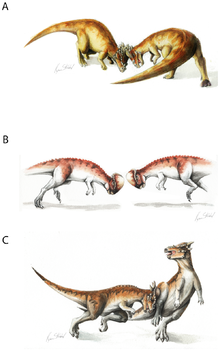
The popular hypothesis among the general public that the skull was used in head-butting, as sort of a dinosaurian battering ram, was first proposed by Colbert (1955), p. [page needed].
Anatomical evidence for combative behavior includes vertebral articulations providing spinal rigidity, and the shape of the back indicating strong neck musculature.
They argued that the dome was "an ephemeral ontogenetic stage", the spongy bone structure could not sustain the blows of combat, and the radial pattern was simply an effect of rapid growth.
Specimens are far less intact; usually only skull caps are recovered, and those found regularly exhibit surface exfoliation and other signs that they were transported long distances by water before fossilization.
The first, occurring before the late Santonian or early Campanian, involved a migration from Asia to North America, most likely by way of the Bering Land Bridge.
The second event occurred before the middle Campanian, and involved a migration back into Asia from North America by a common ancestor of Prenocephale and Tylocephale.
Two species originally reported to be pachycephalosaurs discovered outside this range, Yaverlandia bitholus of England and Majungatholus atopus of Madagascar, have recently been shown to actually be theropods.