Pararhabdodon
Pararhabdodon (meaning "near fluted tooth" in reference to Rhabdodon) is a genus of tsintaosaurin hadrosaurid dinosaur, from the Maastrichtian-age Upper Cretaceous Tremp Group[a] of Spain.
The first remains were discovered from the Sant Romà d’Abella fossil locality and assigned to the genus Rhabdodon, and later named as the distinct species Pararhabdodon isonensis in 1993.
Specimens from other sites, including remains from France, a maxilla previously considered the distinct taxon Koutalisaurus kohlerorum, an additional maxilla from another locality, the material assigned to the genera Blasisaurus and Arenysaurus, and the extensive Basturs Poble bonebed have been considered at different times to belong to the species, but all of these assignments have more recently been questioned.
Excavation of specimens that would later be used to erect Pararhabdodon began in Spring 1985, at the Sant Romà d’Abella (SRA) locality (in the Pyrenees near Isona, Lleida, Spain) of the Talarn Formation[c] in the Tremp Group.
[a][2][3][4] In 1987, Casanovas-Cladellas et al. described remains of an ornithopod from Catalonia, including a cervical vertebra, some partial dorsals, a humerus, and a fragmentary scapula, as Rhabdodon sp.
[5] New remains from this site, excavated in 1990,[4] brought about a reconsideration of the material, and Casanovas-Cladellas and colleagues named it as the new genus and species Pararhabdodon isonense in 1993.
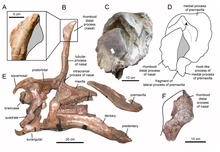
[6] Additional material from the type locality was collected in 1994 - including two maxillae, two dorsal vertebrae, a complete sacrum, two fragmentary ribs, and a partial ischium.
The species name was corrected to isonensis in 1997 by Laurent et al..[7] The new material, most importantly cranial and mandibular elements (those from the skull) led to Casanovas-Cladellas' team re-classifying it as a hadrosaurid as opposed to a basal iguanodont.
[11] In their 1997 paper, Laurent and colleagues referred remains from the Le Bexen site of the uppermost Cretaceous of Aude, southern France, to the genus.
They concluded all the specimens, excepting one fragmentary humerus, were too poorly preserved to allow proper comparison with the holotype material.
[14][15] It was suggested the material likely belong to Koutalisaurus, based on geographic proximity, but this reassignment was abandoned when that genus was recognized as indeterminate.
[11] Near the village Abella de la Conca, in the 1990s, palaeontologist Marc Boada discovered a new site in the Talarn Formation,[c] bearing dinosaur fossils.
One autapomorphy (unique trait) was used to diagnose the taxon as a new species: a very elongated edentulous section on the dentary, which was medially (inwardly) extended.
The authors noted the possibility that future discoveries could lead to synonymization of their taxon with P. isonensis[3] Evidence for this synonymy would later come in a 2009 study, from Prieto-Márquez alongside Jonathan R. Wagner.
As both taxa from the Talarn Formation[c] uniquely shared traits with the Asian genus, the authors decided to treat the two as one species, as maintaining them as provisionally separate would in their eyes be misleading to non-specialists, who would likely not distinguish that the two taxa were being kept separate to be conservative and not due to strong evidence for two hadrosaurs in the area.
[3] Histological analysis indicates the individual was not fully grown, and so the species likely reached similar sizes to North American and Asian relatives such as Corythosaurus and Tsintaosaurus (around 9 metres (30 ft) long[18]).
This is despite Pararhabdodon living on an island, something generally associated with insular dwarfism, a phenomenon exhibited in other European hadrosaurs; Adynomosaurus was cited as a similar example, being around the same size.
Characters of the skeleton supporting this viewpoint were the truncated and rounded anatomy of the articulation of the maxilla to the jugal, the truncated nature of the back of the maxilla itself, the ventral deflection of the front of the dentary (thought the only known dentary was later referred to a new species, Koutalisaurus, and later declared that of indeterminate lambeosaurine[10]), its tall neural spines, and deltopectoral crest of the humerus being distally projected.
[4] The proposal that P. isonensis was the first known European lambeosaurine was soon challenged, however, in 2001 by Jason Head, in a study re-evaluating the status of another species, Eolambia caroljonesa, as a primitive lambeosaur.
The cladogram of Prieto-Márquez et al. (2006) is seen below on the left:[3] Iguanodon bernissartensis Iguanodon atherfieldensis Probactrosaurus gobiensis Ouranosaurus nigeriensis Protohadros byrdi Bactrosaurus johnsoni Gilmoreosaurus mongoliensis Telmatosaurus transsylvanicus Tanius sinensis Pararhabdodon isonensis Hadrosaurinae Lambeosaurinae Equijubus normani Probactrosaurus gobiensis Protohadros byrdi Eolambia caroljonesa Tanius sinensis Bactrosaurus johnsoni Telmatosaurus transsylvanicus Tethyshadros insularis Lophorhothon atopus Hadrosaurus foulkii Edmontosaurus annectens Brachylophosaurus canadensis Aralosaurini Jaxartosaurus aralensis Tsintaosaurus spinorhinus Pararhabdodon isonensis Parasaurolophini Lambeosaurini Prieto-Márquez would return to the issue in 2009 along with Jonathan R. Wagner.
As opposed to European lambeosaurs being spread across different lineages, all members of the subfamily from the continent — including Pararhabdodon — were found to form one monophyletic clade, which was therein named Arenysaurini.