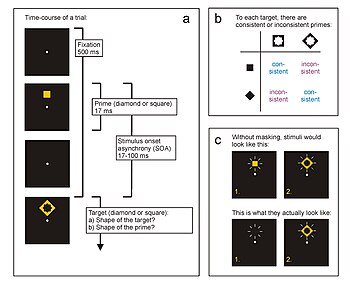
Response priming
In 1962, Fehrer and Raab[1] reported experiments where participants were required to press a single key as quickly as possible upon presentation of a visual target stimulus.
The modern procedure of response priming was developed in the 1980s and 1990s by Peter Wolff, Werner Klotz, Ulrich Ansorge, and Odmar Neumann at the University of Bielefeld, Germany.
[3][4][5][6] The paradigm was developed further in the 1990s by a research team led by Dirk Vorberg at the University of Braunschweig, Germany.
Here, the participant responds as quickly as possible to the shape of the target stimulus (diamond or square) by pressing the assigned response key.
Results from many experiments show that the increase of priming with SOA occurs because the prime has an increasing amount of time to influence the response process before the actual target stimulus can come into play and control the motor response on its own.
This is evident from the time-course of motor activity in the EEG,[8][9][10][11][12] from primed pointing responses,[13][14][15] from measurements of response force,[16] and from simulation studies,[7] which all suggest that motor activation first occurs in the direction specified by the prime, and only then proceeds in the direction specified by the actual target.
Without masking, performance would be nearly perfect; the participant would have little difficulty classifying the prime correctly as a square or a diamond in every trial.
However, when participants had to respond as quickly as possible to the direction of the target, priming effects in all those conditions were virtually identical.
[1][3][4][5][13][14][19][32][33][34][35][36][37] The independence of priming and visual awareness clearly contradicts the traditional notion that effects of unconscious perception merely reflect some residual processing ability under very unfavorable viewing conditions, something that remains after a prime stimulus has been degraded so severely that conscious awareness of it has decreased below some "threshold".
This conception has often led to sharp criticism of research on unconscious or "subliminal" perception,[38][39][40][41] but it is probably wrong on a basic level.
In other words: for a short time and under suitable experimental conditions, visually masked (invisible) stimuli can influence motor responses just as effectively as visible ones.
Sometimes, three stimulus types are employed (prime, mask, target), especially when the prime-target SOA has to be very long.
[33] In one study, chess configurations were presented as primes and targets, and participants had to decide whether the king was in check.
4) was proposed by Odmar Neumann at the University of Bielefeld, Germany, to explain the Fehrer-Raab effect as well as the results of early response priming studies.
[50] The theory assumes that at the outset of a response priming experiment, participants acquire rules of stimulus-response assignment, which quickly become automatized.
This incoming stimulus feature then defines the last missing action parameter (e.g., left vs. right keypress).
However, the conscious representation of the stimuli doesn't play any role for the motor processes in the current experimental trial.
The action trigger account was developed by Wilfried Kunde, Andrea Kiesel, and Joachim Hoffmann at the University of Würzburg, Germany.
Instead, it is assumed that the prime fits a pre-existing action release condition, eliciting the assigned response like a key opening a lock.
In the first step, action triggers are held active in working memory that fit the respective task and are able to elicit a specific motor response.
Based on the instruction, action triggers are set up that automatically elicit the response assigned to the prime or target number.
One important prediction of this theory is that responses can be elicited by primes that meet the trigger conditions but never actually occur as targets.
[52][53] Again, the conscious representation of the stimulus plays no role for motor activation; however, it can lead to a strategic adjustment of response criteria in later trials (e.g., by choosing to respond more slowly to avoid errors).
In sum, this theory can be viewed as expanding on the concept of direct parameter specification by focusing on the exact conditions that lead to response priming.
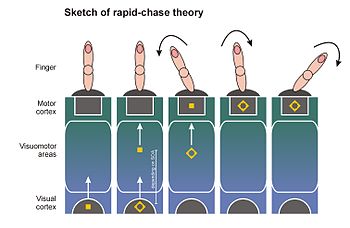
The rapid-chase theory of response priming[12][14][15] was proposed in 2006 by Thomas Schmidt, Silja Niehaus, and Annabel Nagel.
It ties the direct parameter specification model to findings that newly occurring visual stimuli elicit a wave of neuronal activation in the visuomotor system, which spreads rapidly from visual to motor areas of the cortex.
[54][55][56][57] Because the wavefront of activity spreads very fast, Victor Lamme and Pieter Roelfsema from the University of Amsterdam have proposed that this wave starts as a pure feedforward process (feedforward sweep): A cell first reached by the wavefront has to pass on its activity before being able to integrate feedback from other cells.
[29][55] According to rapid-chase theory, both primes and targets elicit feedforward sweeps that traverse the visuomotor system in rapid succession until they reach motor areas of the brain.
[29][55] The most important prediction of rapid-chase theory is that the feedforward sweeps of prime and target signals should occur in strict sequence.