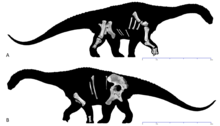
Savannasaurus
Combined with a flexible vertebral column, these traits would have made Savannasaurus better at navigating the muddy ground of the floodplains that it lived on.
Palaeobiogeographic analyses suggest that the ancestors of Savannasaurus and Diamantinasaurus were from either Asia or one of the southern Gondwanan continents; regardless, they likely migrated to Australia through Antarctica around 105–100 million years ago.
In March 2005, the first specimen of Savannasaurus was discovered by David Elliot, founder of Australian Age of Dinosaurs (AODF), along with his wife Judy and their children.
A siltstone concretion containing the specimen was split along gypsum-filled fractures using jackhammers and chisels, with the pieces being marked to facilitate reassembly.
The marks allowed the orientation of the specimen as it was found to be reconstructed, which informed a three-dimensional site map constructed with photogrammetry.
[3] Having been nicknamed "Wade",[2] AODF 660 was formally described and named in 2016 by Stephen Poropat and colleagues in a paper that also described a skull belonging to Diamantinasaurus.
There was a well-developed longitudinal fossa, or excavation, along the side of the vertebra that extends for nearly its entire length, unlike in many titanosaurs where it was faint or absent altogether.
Although titanosaurs were wide-bodied in general, these proportions made Savannasaurus extremely wide-hipped, surpassed only by Opisthocoelicaudia and saltasaurines such as Neuquensaurus.
On the ischium, the iliac peduncle, one of the processes surrounding the acetabulum (hip joint), was only 32% of the bone's length, lower than most other sauropods.
[3] Another distinguishing characteristic of Savannasaurus is the presence of a subtle ridge that separates the front upper portion of the pubis from the rear plate.
[3] For their 2016 description, Poropat and colleagues conducted a phylogenetic analysis to test the affinities of Savannasaurus and the new skull specimen of Diamantinasaurus.
They added these specimens to a dataset created by Philip Mannion (one of the co-authors of the description of Savannasaurus) and colleagues for the 2013 redescription of Lusotitan.
[12] In 2020, Poropat and colleagues noted a number of features that support a position for Savannasaurus outside Lithostrotia: the horizontal transprezygapophyseal laminae, the lack of constriction in the vertebrae of the sacrum, the amphicoelous caudal centra, the pneumatic excavations on the front caudals, the D-shaped sternal plates, the presence of phalanges, and the lack of a longitudinal ridge on the side of the pubis.
At the same time, the lack of hyposphene-hypantrum articulations in the dorsal and caudal vertebrae provides evidence against a more basal (non-specialized or rootward) position, given their presence in many non-titanosaur titanosauriforms and rarity among titanosaurs.
Royo-Torres and colleagues repeated their 2017 analysis after removing a titanosauriform from the Cloverly Formation, Chubutisaurus, and Angolatitan; they found instead that Savannasaurus and Diamantinasaurus were successively basal to a clade consisting of Baotianmansaurus and Dongyanosaurus.
Saltasauridae) Topology B: Royo-Torres et al. (2017)[7] Brachiosauridae Euhelopodidae Sauroposeidon Paluxysaurus Wintonotitan Padillasaurus Dongbeititan Ligabuesaurus Huanghetitan liujiaxiensis Huanghetitan ruyangensis Ruyangosaurus Andesaurus Savannasaurus Diamantinasaurus Diamantinasaurus skull (AODF-836) Dongyangosaurus Baotianmansaurus Xianshanosaurus Daxiatitan Lithostrotia (incl.
[16] Conventional hypotheses suggest that the faunas from the Gondwanan continents (including Australia) would be closely related,[17] but mid-Cretaceous Australian dinosaurs have been found to be more closely related to their Laurasian (northern) counterparts in spite of the fact that Laurasia and Gondwana had already been separated by the Tethys Ocean for a significant span of time.
Incorporating Cretaceous geography suggested ancestral ranges also spanning the Gondwanan continents of South America, Africa, and Indo-Madagascar.
[1] Combining their palaeobiogeographic analysis with time-calibrated phylogenies based on fossil age estimates, Poropat and colleagues inferred that somphospondylans and titanosaurs had attained a wide distribution across the world by the Barremian epoch, 131 to 125 million years ago, if not earlier.
They estimated that the Savannasaurus+Diamantinasaurus and Wintonotitan lineages had separately reached Australia no later than the late Albian epoch, 105 to 100 million years ago, but the exact timing depended on the models that they used.
Yet, Poropat and colleagues also noted traits that would have increased the flexibility of the vertebral column, namely the absence of hyposphene-hypantrum articulations and the presence of prominent excavations (the centroprezygapophyseal fossae) below the transprezygapophyseal laminae.
[1][3] Poropat and colleagues noted that both Savannasaurus and Diamantinasaurus would have lived on a floodplain with clay-rich soil and high rainfall (see § Palaeoecology), which would imply that the ground would occasionally become muddy and hazardous for large sauropods.
They considered it possible that environmental pressures would have selected for individuals capable of navigating muddy ground, and that Savannasaurus might have spent more time near water than other sauropods.
They postulated that saltasaurines might have been subject to similar selective pressures, and so the "typical" titanosaur bodyplan may have been acquired convergently multiple times.
These rocks suggest that Savannasaurus lived on a freshwater floodplain covered in broad, meandering river channels with low-energy waterflow, which was subject to occasional flooding.
However, weather patterns also appeared to have been cyclical on a multi-year basis, similar to the modern Pacific decadal oscillation; this may also have influenced the sporadic flooding.
Other plants included the ferns Phyllopteroides and Tempskya; the liverwort Marchantites; the horsetail Equisetites; the bennettitaleans Otozamites and Ptilophyllum; and the ginkgoalean Ginkgo.
[26] Contemporary dinosaurs included the sauropods Diamantinasaurus matildae and Wintonotitan wattsi, the megaraptoran theropod Australovenator wintonensis, and indeterminate ankylosaurians and hypsilophodonts.
Additional vertebrates known from the Winton Formation are chelid turtles; an indeterminate varanoid lizard;[30] the lungfish Metaceratodus ellioti and M. wollastoni; and ray-finned fish including Cladocyclus geddesi.
[31] Invertebrates were dominated by the bivalves Alathyria jaqueti, Hyridella goondiwindiensis, Megalovirgus wintonensis, and Prohyria macmichaeli, but also included the gastropod Melanoides, dragonflies, scorpionflies, and oribatid mites known from trace fossils found in silicified wood.