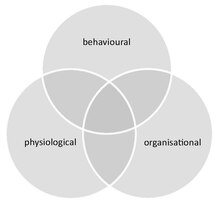
Social immunity
Empirical and theoretical work in social immunity continues to reveal not only new mechanisms of protection but also implications for understanding of the evolution of group living and polyandry.
[1] Upon exposure to a parasite, group members must both evaluate the threat it poses and the current level of colony infection in order to respond appropriately.
[6] For a parasite to succeed in infecting multiple members of an insect group, it must complete three key tasks: Mechanisms of social immunity are thus often categorized by which step(s) they hinder and/or block.
[2] In eusocial insects, the most frequent defence against parasite uptake into the nest is to prevent infection during and/or after foraging,[1] and a wide range of active and prophylactic mechanisms have evolved to this end.
Allogrooming is found in many animals—for example primates frequently groom others, a behaviour which likely evolved for its hygienic function but has now been co-opted for its additional role in social bonding.
Corsican blue tits (Parus caeruleus) prophylactically line their nest with aromatic plants (such as Achillea ligustica, Helichrysum italicum and Lavandula stoechas) to ward off mosquitoes and other blood-sucking ornithophillous (bird-targeting) insects.
[64] After the broader definition of social immunity by Cotter and Kilner, numerous examples of social immune behaviours within animal families can be given: túngara frogs (Engystomops pustulosus) create 'foam nests' during breeding in which embryogenesis occurs; these foam nests are imbued with ranaspumin proteins which provide defence against microbial attack and act as a detergent.
The three-spined stickleback (Gasterosteus aculeatus), grass goby (Zosterisessor ophiocephalus), fringed darter (Etheostoma crossopterum) and two species of blenny also use chemical strategies to defend their eggs from microbes.
[65] An analogy can be drawn here with the social fever in bees described above: a subset of individuals in a population change their behaviour and in doing so provide population-wide resistance.
[37] Cotter et al. went on to show the costliness of this social immune response-by providing females with microbe-infested carcasses, they found that the upregulation of antibacterial activity that followed led to a 16% decrease in lifetime reproductive output.
[67] This significant reduction in fitness, due to both increased mortality and age-related dropoff in fecundity, explains why the antibacterial activity of the exudate is only induced and not present constitutively.
Example analogies from Cremer's paper are: Other similarities include the immunological memory of the adaptive immune system in vertebrates and the observation that a similar collective memory (operating with a yet-to-be-explained mechanism) occurs in some insect societies e.g. individual Z. angusticollis survive M. anisopliae infections significantly more when they have been in contact with a previously infected conspecific, a 'social transfer of immunity' or 'social vaccination'.
[4][72] Transplant rejection caused by non-self major histocompatibility complexes is frequently thought to be a byproduct with no evolutionary function, however Cremer cites cases (such as the colonial star ascidian (Botryllus schlosseri)) where recognizing foreign cells may have evolved as an adaptation - if so, then this could be analogous to the self-recognition systems in social insects which prevent brood parasitism and the worker policing behaviours which suppress 'social tumours'.
[1] More studies on presocial taxa would allow for phyletic analyses to recover the actual path of evolution that different mechanisms of social immunity took.
[1] The origin of polyandry in nature and its adaptive value is a subject of ongoing controversy in evolutionary biology, partly due to the seemingly numerous costs it places on females - additional energetic and temporal allocation to reproduction, increased risk of predation, increased risk of sexually transmitted diseases and increased risk of physical harm caused by copulation/sexual coercion – for eusocial insects, the effects polyandry has on the colony member's coefficient of relatedness is also important, as reducing the relatedness of workers limits the power of kin selection to maintain the ultracooperative behaviours which are vital to a colonies' success.
For example, organisational defences, such as an altered interaction network that influences disease spread, emerge from chemical and behavioural processes.
[91] As social organisms are often densely aggregated and exhibit high levels of interaction, pathogens can more easily spread from infectious to susceptible individuals.
For example, many social insects nest and forage in habitats that are rich in pathogens, such as soil or rotting wood, exposing them to a plethora of microparasites, e.g. fungi, bacteria, viruses and macroparasites, e.g. mites and nematodes.
[93] In addition, shared food resources, such as flowers, can act as disease hubs for social insect pollinators, promoting both interspecific and intraspecific pathogen transmission.
All of these factors combined can therefore contribute to rapid disease spread following an outbreak, and, if transmission is not controlled, an epizootic (an animal epidemic) may result.
Social insects have evolved an array of sanitary behaviours to keep their nests clean, thereby reducing the probability of parasite establishment and spread within the colony.
Secretions from the metapleural glands of ants and volatile chemical components produced by termites have been shown to inhibit fungal germination and growth.
[113] For xeric-living ants, placing waste outside will tend to inhibit infectious material, as microbes are usually killed under hot, dry conditions.
This "social fever" is performed before symptoms of the disease are expressed and can therefore be viewed as a preventative measure to avoid chalk brood outbreaks in the colony.
[123] In addition to grooming, social insects can apply host- and symbiont-derived antimicrobial compounds to themselves and each other to inhibit pathogen growth or germination.
[126] Cannibalism of infected nest mates is an effective behaviour in termites, as ingested infectious material is destroyed by antimicrobial enzymes present in their guts.
[110] Some fungal pathogens (e.g. Ophiocordyceps, Pandora) manipulate their ant hosts into leaving the nest and climbing plant stems surrounding the colony.
[141] In all cases, social contact with pathogen-exposed individuals promoted reduced susceptibility in their nestmates (increased survival), upon subsequent exposure to the same pathogen.
In the ant-fungus [140] and termite-fungus [142] systems, social immunisation was shown to be caused by the transfer of fungal conidia during allogrooming, from the exposed insects to nestmates performing grooming.
This contamination resulted in low-level infections of the fungus in the nestmates, which stimulated their immune system, and protected them against subsequent lethal exposures to the same pathogen.