Hippocampus anatomy
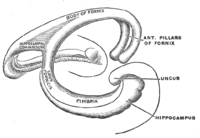
In humans and other primates, the portion of the hippocampus near the base of the temporal lobe is much broader than the part at the top.
This led to its description of cornu ammonis meaning Amun's horn, whose initials were used to name the subfields CA1-CA4 that make up the structure of the hippocampus.
Its curved layers are of different cell densities and varying degrees of axons in the subfields.
Starting at the dentate gyrus and working inward along the S-curve of the hippocampus means traversing a series of narrow zones.
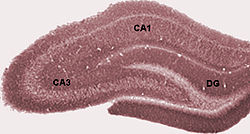
The first of these, the dentate gyrus (DG), is actually a separate structure, a tightly packed layer of small granule cells wrapped around the end of the hippocampus proper, forming a pointed wedge in some cross-sections, a semicircle in others.
Next come a series of Cornu Ammonis areas: first CA4 (which underlies the dentate gyrus), then CA3, then a very small zone called CA2, then CA1.
The CA areas are all filled with densely packed pyramidal cells similar to those found in the neocortex.
The perforant path-to-dentate gyrus-to-CA3-to-CA1 was called the trisynaptic circuit by Per Andersen, who noted that thin slices could be cut out of the hippocampus perpendicular to its long axis, in a way that preserves all of these connections.
This observation was the basis of his lamellar hypothesis, which proposed that the hippocampus can be thought of as a series of parallel strips, operating in a functionally independent way.
[5] The lamellar concept is still sometimes considered to be a useful organizing principle, but more recent data, showing extensive longitudinal connections within the hippocampal system, have required it to be substantially modified.
[7][8][9] Region CA1 receives input from the CA3 subfield, EC layer III and the nucleus reuniens of the thalamus (which project only to the terminal apical dendritic tufts in the stratum lacunosum-moleculare).
In turn, CA1 projects to the subiculum as well as sending information along the aforementioned output paths of the hippocampus.
[13][14] Different parts of this system are given different names: At the circuit level, the alveus contains axonal fibers from the DG and from pyramidal neurons of CA3, CA2, CA1 and the subiculum (CA1 ▶ subiculum and CA1 ▶ entorhinal projections) that collect in the temporal hippocampus to form the fimbria/fornix, one of the major outputs of the hippocampus.
[12] As a rule of thumb, one could say that each cytoarchitectonic field that contributes to the commissural projection also has a parallel associational fiber that terminates in the ipsilateral hippocampus.
However, associational/commissural fibers, like CA2 Pyramidal cell associational projections, span mainly longitudinally (dorsoventrally) along the hippocampus.
[34] Although excitatory cells are the main contributors to commissural pathways, a GABAergic component has been reported among their terminals which were traced back to hilus as origin.
The CA regions are also structured depthwise in clearly defined strata (or layers): The dentate gyrus is composed of a similar series of strata: An up-to-date knowledge base of hippocampal formation neuronal types, their biomarker profile, active and passive electrophysiological parameters, and connectivity is supported at the Hippocampome website.