Tarsophlebiidae
They are characterized by the basally open discoidal cell in both pairs of wings, very long legs, paddle-shaped male cerci, and a hypertrophied ovipositor in females.
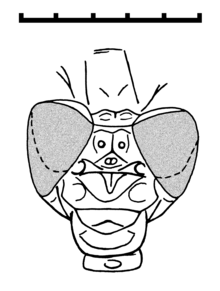
[1] The head is similar to that of Recent Gomphidae with two large and globular compound eyes that are distinctly separated, but closer together than in damselflies (Zygoptera).
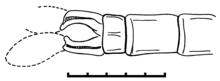
The description of calopterygoid-like appendages of Tarsophlebia eximia, with apparently two pairs of claspers,[2][4] was based on misinterpretations due to artifacts of preservation.
The broken double-barrelled petioles of the two cerci have been commonly misinterpreted as two pairs of claspers, while the distal plates have been overlooked or regarded as artifacts.
[5] Tarsophlebiopsis mayi was recognized as valid by most authors until the most recent revision of the family by Fleck et al. (2004),[6] who demonstrated that the holotype of this taxon most probably is only an aberrant specimen of Tarsophlebia eximia.
The body characters "distinctly prolonged legs, with very long tarsi" and "male cerci with paddle-like distal expansions" are known from one species of the genus Tarsophlebia (T. eximia) and Turanophlebia (T. vitimensis) respectively, and thus belonged to the common ground plan of all Tarsophlebiidae.
[1] Bechly (1996) and Nel et al. (2004: Appendix 2) proposed a long list of putative synapomorphies that demonstrate a closer relationship of Tarsophlebiidae with crown group Odonata: presence of a costal triangle as broad and strong sclerotisation of the basal costal margin; the distal discoidal vein MAb (= distal side of discoidal cell) and the subdiscoidal vein (origin of the CuA on MP) are aligned and dorsally enforced by a strong sclerotisation, so that this structure appears to cross the vein MP and the concave fold along this vein (formation of a “discal brace” sensu Carle 1982); this discal brace is aligned with the arculus in the ground plan (only retained in some taxa with an open discoidal cell and in the forewings of Epiophlebia); the midfork (first fork of RP and base of IR2) is shifted basally, with the RP3/4 generally arising basal of the subnodus (reversed in some Coenagrionoidea) and RP2 arising close to the subnodus (in the ground plan); more derived type of nodus, with a kink in ScP; the oblique basal brace (still present in Protanisoptera and Protozygoptera) is transformed into a transverse “basal bracket” Ax0 which looks like a primary antenodal crossvein; presence of two strong primary antenodal crossveins Ax1 and Ax2 (also present in some Protozygoptera, e.g. specimen PIN 1/276 from the Upper Permian of Russia); pterostigma distinctly braced by an oblique postsubnodal crossvein beneath the basal margin of the pterostigma; presence of a tracheated lestine oblique vein 'O' between RP2 and IR2 (secondarily absent in some taxa); in the median space (= basal space) the convex vestige of the Media-stem (“vestigial CuA” sensu Fraser 1957) is suppressed since it is fused with the cubital stem to a common medio-cubital-stem (the alleged presence of this vestige in the holotype of Tarsophlebiopsis is a teratology or an artefact of preservation), convergent to some Protanisoptera, Triadophlebiomorpha, and Protozygoptera.
[4] These features are the basally open discoidal cell in the hindwing (instead of closed) which implies an incomplete arculus, the (meanwhile disputed) presence of four tarsomeres of equal length (instead of only three), and the very primitive condition of the male secondary genital apparatus (viz ligula orimentary; vesicula spermalis still very short and flat with a very wide porus) without any intromittent organ.
Bechly therefore considered the similarities of Tarsophlebiidae and Epiprocta mentioned by Nel et al. (1993), viz the less separated and relatively large eyes, the presence of two cephalic sutures, and the small leg spines (also present in Meganisoptera[3]), as symplesiomorphies.
However, based on a cladistic study of 14 characters Fleck et al. (2004) again suggested that Tarsophlebiidae might rather be the sister group of the clade Epiprocta that includes Epiophlebiidae and Anisoptera.
Nevertheless, this result has a very low statistical support and might as well be an artefact of the parsimony computer algorithm, because none of the 14 characters represents an unambiguous synapomorphy for Tarsophlebiidae and Epiprocta.
The fossil record of this family ranges from Upper Jurassic of Kazakhstan and Germany to the Lower Cretaceous of England, Transbaikalia, Mongolia and China.