Valosin-containing protein
[9] It has become evident that the interplays between ubiquitin and VCP cofactors are critical for many of the proposed functions, although the precise role of these interactions remains to be elucidated.
[9] In mammalian cells, VCP is predominantly localized to the cytoplasm, and a significant fraction is associated to membranes of cellular organelles such as the endoplasmic reticulum (ER), Golgi, mitochondria, and endosomes.
For example, VCP mutant proteins carrying single point mutations found in patients with multisystem proteinopathy (MSP; formerly known as IBMPFD (inclusion body myopathy associated with Paget disease of the bone and frontotemporal dementia)) (see below) have 2-3 fold increase in ATPase activity.
Many of these proteins serve as adaptors that link VCP to a particular subcellular compartment to function in a specific cellular pathway.
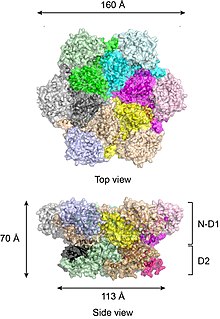
The N-domain consists of two sub-domains of roughly equal size: the N-terminal double Y-barrel and a C-terminal b-barrel (Figure 3).
VCP is also required to release defective translation products stalled on ribosome in a process termed ribosome-associated degradation.
[56] VCP also functions broadly in eukaryotic nucleus by releasing protein molecules from chromatins in a manner analogous to that in ERAD.
These substrates link VCP function to gene transcription, DNA replication and repair, and cell cycle progression.
Biochemical and genetic studies have also implicated VCP in fusion of vesicles that lead to the formation of Golgi apparatus at the end of mitosis.
Recent studies also suggest that VCP may regulate vesicle trafficking from plasma membrane to the lysosome, a process termed endocytosis.
[56] Antibody fragment-based inhibitors have been developed by a team led by Arkin to inhibit the interaction between p97 and p47, selectively modulating the Golgi reassembly process.
[59] Mutations in VCP were first reported to cause a syndrome characterized by frontotemporal dementia, inclusion body myopathy, and Paget's disease of the bone by Virginia Kimonis in 2004.
[60] In 2010, mutations in VCP were also found to be a cause of amyotrophic lateral sclerosis by Bryan Traynor and Adriano Chiò.
[61] This discovery was notable as it represented an initial genetic link between two disparate neurological diseases, amyotrophic lateral sclerosis and frontotemporal dementia.
In 2020, Edward Lee described a distinct hypomorphic mutation in VCP associated with vacuolar tauopathy, a unique subtype of frontotemporal lobar degeneration with tau inclusions.
[63] Although MSP is rare, growing interest in this syndrome derives from the molecular insights the condition provides into the etiological relationship between common age-related degenerative diseases of muscle, bone and brain.
The first p97 inhibitor Eeyarestatin (EerI) was discovered by screening and characterizing compounds that inhibit the degradation of a fluorescence-labeled ERAD substrate.
[68][69][70] More recently, a potent and specific VCP inhibitor CB-5083 has been developed, which demonstrates promising anti-cancer activities in mouse xenograft tumor models.