Variant surface glycoprotein
[1] VSG allows the trypanosomatid parasites to evade the mammalian host's immune system by extensive antigenic variation.
[citation needed] For this reason, these proteins are highly immunogenic and an immune response raised against a specific VSG coat will rapidly kill trypanosomes expressing this variant.
The parasite has a large cellular repertoire of antigenically distinct VSGs (~1500/2000[citation needed] complete and partial (pseudogenes)) located in telomeric and subtelomeric arrays (on megabase chromosomes or minichromosomes).
This coat replacement process is critical for the survival of recently switched cells because initial VSGs remain targets for the escalating host Ab response.
VSG half-life measurements suggest that initial VSGs may persist on the surface of genetically switched trypanosomes for several days.
The repetition of this process prevents the extinction of the infecting trypanosome population, allowing chronic persistence of parasites in the host and enhancing opportunities for transmission.
[citation needed] The properties of the VSG coat that enable immune evasion are: The VSGs from T. brucei are attached to the plasma membrane via a covalent attachment to two glycosyl-phosphatidylinositol (GPI) anchors (one per monomer),[7] which directs its forward-trafficking from the ER to the flagellar pocket for incorporation into the membrane, as predicted by the GPI valence hypothesis.
[8][9] VSGs are replaced by an equally dense coat of procyclins when the parasite differentiates into the procyclic form in the tsetse fly midgut.
The formation of mosaic VSGs allows the (partial) expression of pseudogene VSGs, which can constitute the major portion of the VSG archive, and can contribute directly to antigenic variation, vastly increasing the trypanosome's capacity for immune evasion and posing a major problem for vaccine development.
These are generally silent, typically with omitted sections or premature stop codons, but are important in the evolution of new VSG genes.
[19] Trypanosoma have a simple, polarized membrane transport system consisting of a single ER, lysosome, and Golgi apparatus.
Importantly, following incorporation into the cellular membrane, VSG may later be recycled through the flagellar pocket and sorted back to the cell surface.
VSG is not turned over by lysosomal or proteasomal canonical degradation pathways,[21] but is instead lost from the cell by specific cleavage of its GPI anchor by GPI-specific PLC.
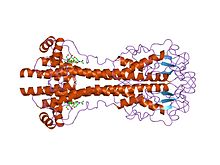
VSG genes are hugely variable at the sequence (primary) level, but variants are thought to have strongly conserved structural (tertiary) features, based on two determined 3-dimensional structures[22] and conservation of 2-dimensional sequence motifs (descending and ascending alpha-helices that make up the dimerization interface), allowing them to perform a similar shielding function.
Reiteration of this process prevents extinction of the infecting trypanosome population, allowing chronic persistence of parasites in the host, enhancing opportunities for transmission.
[27] A VSG gene from Trypanosoma evansi, a parasite that causes a form of surra in animals, has been cloned in Escherichia coli.
The animals immunized with whole cell lysate or recombinant protein show similar antibody reactions in ELISA (enzyme-linked immunosorbent assay) and CATT (card agglutination test for trypanosomiasis).
[28] The variable surface glycoprotein RoTat 1.2 PCR can be used as a specific diagnostic tool for the detection of T. evansi infections.