Wood-decay fungus
Some species of wood-decay fungi attack dead wood, such as brown rot, and some, such as Armillaria (honey fungus), are parasitic and colonize living trees.
[4] Because hydrogen peroxide is a small molecule, it can diffuse rapidly through the wood, leading to a decay that is not confined to the direct surroundings of the fungal hyphae.
[citation needed] Soft-rot fungi are able to colonise conditions that are normally too hot, cold or wet for brown- or white-rot to inhabit.
[17] White-rot fungi show strong participation in interspecific competition, culminating in the evolution of lignin catabolism specificity.
The current and future applications of white-rot fungi as a potential component of mycoremediation merit greater study of these saprotrophs.
[18] Compared to other saprotrophs, white-rot fungi possess the specialized ability to cleave lignin into smaller, more processable molecules.
The relative abundance of phenylpropane alkyl side chains of lignin characteristically decreases when decayed by white-rot fungi.
[21] PODs developed in the common ancestor of white-rot, brown-rot and mycorrhizal fungi but these enzyme families have undergone secondary loss or contraction in the latter two groups.
LiPs and VPs are specific to heme product architecture allowing direct oxidation of benzene groups regardless of linkages.
As a result, LiP and VP enzymes create a tryptophan radical on their protein surface which allows long-range electron transfer from the aromatic substrate to the activated cofactor.
While the general pathway is currently unknown, research supports the existence of a continuum of features that separate the two fungal types rather than distinct categories.
Another method involves endoglucanases hydrolyzing cellulose at random points before cellobiohydrolases cleave the chains, resulting in cellobiose.
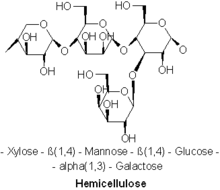
The prevalent hemicellulose found in soft wood trees is Galactoglucomannan, a molecule made up of b-1,4-linked D-mannopyranose and D-glucopyranose units.
[29] The availability of non white-rot fungi to catabolize cellulose and hemicellulose results in the creation of interspecific competition for access to these resources.
[citation needed] Since white-rot fungi aren't the only saprotrophs capable of accessing cellulose and hemicellulose, competition ensues.
Research further highlighted the importance of environmental factors including temperature, water potential, and invertebrate interactions in influencing competition.
Though the mechanism is unknown, researchers suggested that white-rot fungi may utilize lignin decomposing enzymes, hydroxyl radicals, and aryl alcohols to create a toxic environment.
Results suggested that white-rot saprotrophs were the common ancestors of brown-rot fungi and ectomycorrhiza (ECM), but that in the latter two groups genes coding for PODs were lost.
One of the major findings was that ancestral versatile peroxidase (AVP) was not capable of functioning efficiently at low pH, a characteristic associated with modern LiPs.
[37] Early peroxidases were unable to directly degrade lignin and relied on metal cations to separate phenol groups.
White-rot fungi have been determined to degrade chlorinated aromatic hydrocarbons (CAHs), DDT, lindane, polycyclic aromatic hydrocarbons, polychlorinated biphenyls, PCP, polychlorinated dibenzo(p)dioxins, and azo dyes when studied in Phanerochaete chrysosporium, Trametes versicolor, Bjerkandere adusta, and Pleurotus ostreatus.
A special way of giving grown wood an unusual structure is to infect it with a parasitic fungus by storing it in a humid environment (fungal decay).
In order to stabilise the wood structure weakened by the fungus, resins or plastics are usually introduced into the material by special vacuum processes.
[42] A special icing process applied to beech wood leads to results similar to those obtained with fungal decay.
[43][44] Natural durability is the inherent capability of wood to tolerate and resist fungal decay and insect attacks, such as woodboring beetles and termites, and marine organisms.
[47] Different chemicals have been isolated from the heartwood of naturally rot-resistant trees and have shown to be protectants, including polyphenols, lignans (e.g. gmelinol, plicatic acid), flavonoids (e.g. mesquitol), tropolones (e.g. hinokitiol and other thujaplicins), sesquiterpenoids (e.g.
[48][49] The natural durability varies between tree species, geographic regions, environmental conditions, growth stage, and increases with the age.
[51] Tree species that have significant natural durability include Lagarostrobos franklinii (Huon pine), Intsia bijuga (ipil), some Eucalyptus species (ironbark), Podocarpus totara (totara), Vitex lucens (puriri), Agathis australis (kauri), and trees of the Cupressaceae family, such as Chamaecyparis obtusa (Hinoki cypress), Thuja plicata (Western red cedar), Thujopsis dolabrata (Hinoki asunaro), Juniperus cedrus (Canary Islands juniper), Cedrus atlantica (Atlas cedar), Chamaecyparis taiwanensis (Taiwan cypress), among others.
[citation needed] A wide selection of timber preservation has been developed to give the wood an improved durability and to protect it from decay.
[59] Water contamination with arsenic and its compounds is a serious public health issue, and their release to the environment and soil pollution is another environmental problem.