Astrocyte
They perform many functions, including biochemical control of endothelial cells that form the blood–brain barrier,[1] provision of nutrients to the nervous tissue, maintenance of extracellular ion balance, regulation of cerebral blood flow, and a role in the repair and scarring process of the brain and spinal cord following infection and traumatic injuries.
[5] Research since the mid-1990s has shown that astrocytes propagate intercellular Ca2+ waves over long distances in response to stimulation, and, similar to neurons, release transmitters (called gliotransmitters) in a Ca2+-dependent manner.
[8] Astrocytes are classically identified using histological analysis; many of these cells express the intermediate filament glial fibrillary acidic protein (GFAP).
[10] The protoplasmic glia are the most prevalent and are found in grey matter tissue, possess a larger quantity of organelles, and exhibit short and highly branched tertiary processes.
This is supported by 3D neuropil reconstructions indicating similar mitochondrial densities in both cell types, as well as cell-specific transcriptomic and proteomic data, and tricarboxylic acid cycle rates.
The resultant patterning along the neuraxis leads to segmentation of the neuroepithelium into progenitor domains (p0, p1 p2, p3 and pMN) for distinct neuron types in the developing spinal cord.
Astrocytes help form the physical structure of the brain, and are thought to play a number of active roles, including the secretion or absorption of neural transmitters and maintenance of the blood–brain barrier.
Because this influx is directly caused by an increase in blood flow to the brain, calcium waves are said to be a kind of hemodynamic response function.
The close association between astrocytes and presynaptic and postsynaptic terminals as well as their ability to integrate synaptic activity and release neuromodulators has been termed the tripartite synapse.
A 2023 study suggested astrocytes, previously underexplored brain cells, could be key to extending wakefulness without negative effects on cognition and health.
Anaplastic astrocytomas recur more frequently than lower grade tumors because their tendency to spread into surrounding tissue makes them difficult to completely remove surgically.
Pain processing is no longer seen as a repetitive relay of signals from body to brain, but as a complex system that can be up- and down-regulated by a number of different factors.
[51][61][62] In the past, hyperalgesia was thought to be modulated by the release of substance P and excitatory amino acids (EAA), such as glutamate, from the presynaptic afferent nerve terminals in the spinal cord dorsal horn.
These factors increase the responsiveness of the dorsal horn pain-projection neurons to ensuing stimuli, termed "spinal sensitization", thus amplifying the pain impulse to the brain.
Release of glutamate, substance P, and calcitonin gene-related peptide (CGRP) mediates NMDAR activation (originally silent because it is plugged by Mg2+), thus aiding in depolarization of the postsynaptic pain-transmitting neurons (PTN).
In addition, activation of IP3 signaling and MAPKs (mitogen-activated protein kinases) such as ERK and JNK, bring about an increase in the synthesis of inflammatory factors that alter glutamate transporter function.
As noxious pain is sustained, spinal sensitization creates transcriptional changes in the neurons of the dorsal horn that lead to altered function for extended periods.
Mobilization of Ca2+ from internal stores results from persistent synaptic activity and leads to the release of glutamate, ATP, tumor necrosis factor-α (TNF-α), interleukin 1β (IL-1β), IL-6, nitric oxide (NO), and prostaglandin E2 (PGE2).
In response to nerve damage, heat shock proteins (HSP) are released and can bind to their respective TLRs, leading to further activation.
They can be found throughout the brain, but are by far the most abundant in the olfactory bulbs, medial habenula, dentate gyrus of the hippocampus, arcuate nucleus of the hypothalamus, and in the dorsal medulla, just beneath the area postrema.
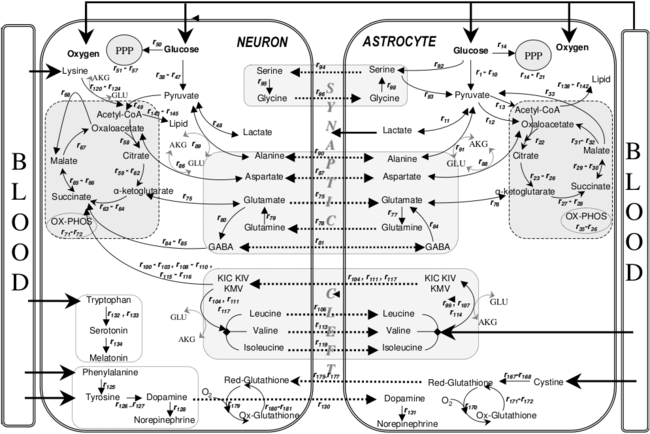
Recent data have shown that astrocytes, but not neurons, possess the mitochondrial enzymes needed to metabolize fatty acids, and that the resulting oxidative stress can damage mitochondria.
For example, GP astrocytes are in close contact with neurons that make a neurotransmitter called dopamine in both the rat and human hypothalamus.
The activity of dopaminergic neurons declines during aging, leading to elevations in blood levels of prolactin that can provoke breast cancer.
FABP7+ astrocytes are in close contact with neurons in the arcuate nucleus of the hypothalamus that are responsive to a hormone called leptin that is produced by fat cells.
Mitochondrial damage in these astrocytes could thus alter the function of leptin-sensitive neurons and could contribute to an aging-associated dysregulation of feeding and body weight.
A single infusion of a protein called fibroblast growth factor-1 into the hypothalamus has been shown to permanently normalize blood glucose levels in diabetic rodents.
They did an experiment to attempt to repair trauma to the Central Nervous System of an adult rat by replacing the glial cells.
So, with the bone protein and human glial cells combined, they promoted significant recovery of conscious foot placement, axonal growth, and obvious increases in neuronal survival in the spinal cord laminae.
[90] In a study published in a 2011 issue of Nature Biotechnology[91] a group of researchers from the University of Wisconsin reports that it has been able to direct embryonic and induced human stem cells to become astrocytes.
More specifically, when astrocytes became reactive they unleash the pathological effects of amyloid-beta on downstream tau phosphorylation and deposition, which very likely will lead to cognitive deterioration.