Brain stimulation reward
This activation is considered to be more direct than rewards produced by natural stimuli, as those signals generally travel through the more indirect peripheral nerves.
ICSS typically involves subjects with permanent electrode implants in one of several regions of the brain known to produce BSR when stimulated.
ICSS has been utilized as a means to gauge addiction liability for drugs of many classes, including those that act on monoaminergic, opioid, and cholinergic neurotransmission.
[6] In 1953, James Olds and Peter Milner, of McGill University, observed that rats preferred to return to the region of the test apparatus where they received direct electrical stimulation to the septal area of the brain.
[1] The natural drives that motivate and shape behavior reach the reward circuitry trans-synaptically through the peripheral senses of sight, sound, taste, smell, or touch.
[2] For this reason, electrical brain stimulation provides a tool for identifying the reward circuitry within the central nervous system with some degree of anatomical and neurochemical specificity.
Both of these effects lead to diminished response rate for BSR in the early trials of a series; however, experiments have also shown that extinguished behavior can be quickly reinstated by a priming stimulation that refreshes the short-term association involved in reward expectancy.
Rats will perform lever-pressing at rates of several thousand responses per hour for days in exchange for direct electrical stimulation of the lateral hypothalamus.
Addiction is a chronic brain disorder consisting of compulsive drug-taking and seeking that is maintained despite detrimental effects on various aspects of life including health, relationships, and work.
[2][15] Similar to self-administration behavior, responding for intracranial brain stimulation has a highly compulsive component characteristic of an addicted state.
Sites along the length of the medial forebrain bundle (MFB) through the lateral and posterior hypothalamus, the ventral tegmental area (VTA), and into the pons are associated with the strongest reward effects of stimulation.
[3] MFB is not the sole anatomical substrate responsible for reinforcing brain stimulation; however, it is the main tract for the ascending dopamine fibers, and it functions to relay information from the VTA to the nucleus accumbens.
The nucleus accumbens is located in the ventral striatum and integrates information from cortical and limbic brain structures to mediate behaviors the reinforce reward.
[3] BSR has been shown to result in the release of dopamine within the nucleus accumbens, which also occurs in response to natural rewards such as food or sex.
[1] Electrophysiological data suggest stimulation of the MFB or VTA does not directly activate dopaminergic neurons in the mesolimbic reward pathway.
Excitatory, cholinergic inputs to the VTA are thought to play a role in this indirect activation, but the neuroanatomical components of this circuit have yet to be fully characterized.
[5][6] Since the initial demonstration of BSR by Olds and Milner, experiments in rodents record ICSS responding to quantify motivation to receive stimulation.
The first portion of an ICSS experiment involves training subjects to respond for stimulation using a fixed-ratio 1 (FR-1) reinforcement schedule (1 response = 1 reward).
In experiments involving rats, subjects are trained to press a lever for stimulation, and the rate of lever-pressing is typically the dependent variable.
Subjects with imperfect electrode placement require a higher simulation amplitude to activate the reward circuitry and produce ICSS responding.
In an experimental condition, if the MCR falls below 100% at the highest stimulation frequencies, it is thought to reflect an impacted capability or motivation to respond, potentially induced by a drug with sedative or aversive properties.
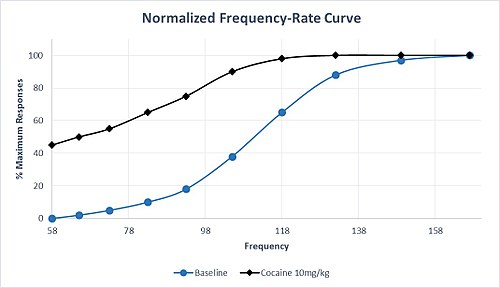
[20] Sensitivity of the neural circuitry to the rewarding properties of stimulation is assessed by analyzing left- or right-shifts in the M50, or the frequency at which 50% of the maximum number of responses was recorded.
An increase in the M50 indicates that a greater stimulation frequency was required to reach 50% of the MCR, and the reward circuitry has been desensitized by the experimental manipulation.
The neuronal axons of the mesolimbic dopamine system, a key component of the reward circuitry, generally have high thresholds for stimulation.
Amphetamine and cocaine share this selectivity profile, and administration of these drugs generally results in a left-shift in M50 and θ0, indicating sensitization of the reward circuitry and high abuse potential.
[6] Drugs with more balanced selectivity for dopamine/norepinephrine and serotonin transmission, such as 3,4-methylenedioxy-methamphetamine (MDMA), tend to be less addictive, and they have mixed effects on ICSS responding depending on dose and stimulation frequency.
Drugs that act on GABAA receptors, which are ligand-gated ion channels, are more widely studied, as they generally produce more robust effects on sedation and anxiety, and they are commonly prescribed for therapeutic uses.
Ethanol influences GABA receptor activity, and has been found to moderately facilitate ICSS,[6] despite older publications suggesting these findings are inconsistent.
ICSS studies of BSR have proven to be a robust measure of reward sensitivity, and have potential to help assess the abuse liability of various future therapeutics.
[31][32][33] Drugs found to prevent ICSS facilitation have potential to be developed and therapeutically implemented to reduce the risk of addictive disorders in a clinical setting.