Calcium sparks
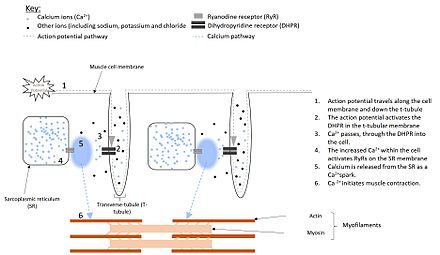
[1] This release occurs through an ion channel within the membrane of the SR, known as a ryanodine receptor (RyR), which opens upon activation.
An increase in Ca2+ within the SR is thought to bind to Ca2+ sensitive sites on the inside of the RyR causing the channel to open.
As well as this, a protein called calsequestrin (found within the SR) detaches from the RyR, when calcium concentration is too high, again allowing the channel to open (see sarcoplasmic reticulum for more details).
This is thought to be due to the calsequestrin binding more strongly to the RyR, preventing it from opening and decreasing the likelihood of a spontaneous spark.
[15] An increase in Ca2+ concentration within the cell or the production of a larger spark, can lead to a large enough calcium released that the neighbouring cluster can be activated by the first.
This is known as spark-induced spark activation and can lead to a Ca2+ wave of calcium release spreading across the cell.
This is because, the Ca2+ that enters the cell via the DHPR in response to the action potential, stimulates both muscle contraction and calcium release from the SR.
[20] However, due to the large numbers of RyRs and DHPRs in a single cell, this theory seems to be unrealistic, as there is a very small probability that they would all close together at exactly the same time.
[18] This theory suggests that after activation of the RyR and the subsequent release of Ca2+, the channel closes briefly to recover.
[21] Spontaneous Ca2+ sparks were discovered in cardiac muscle cells, of rats, in 1992 by Peace Cheng and Mark B. Cannell in Jon Lederer's laboratory at the University of Maryland, Baltimore, U.S.A.
However they were quickly recognised as being of fundamental importance to muscle physiology, playing a huge role in excitation-contraction coupling.
Ca2+ “sparks” were so called because of the spontaneous, localised nature of the Ca2+ release as well as the fact that they are the initiation event of excitation-contraction coupling.