Chromosomal crossover
It is one of the final phases of genetic recombination, which occurs in the pachytene stage of prophase I of meiosis during a process called synapsis.
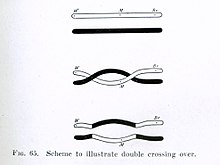
Morgan immediately saw the great importance of Janssens' cytological interpretation of chiasmata to the experimental results of his research on the heredity of Drosophila.
[6][7] When Hotta et al. in 1977 compared meiotic crossing-over (recombination) in lily and mouse they concluded that diverse eukaryotes share a common pattern.
McClintock used the prophase and metaphase stages of mitosis to describe the morphology of corn's chromosomes, and later showed the first ever cytological demonstration of crossing over in meiosis.
Working with student Harriet Creighton, McClintock also made significant contributions to the early understanding of codependency of linked genes.
[12] Several other genes in D. melanogaster have been linked as well to both processes, by showing that mutants at these specific loci cannot undergo DNA repair or crossing over.
[citation needed] This large group of conserved genes between processes supports the theory of a close evolutionary relationship.
In an experiment using radiation hybrid mapping on wheat's (Triticum aestivum L.) 3B chromosome, crossing over and DNA repair were found to occur predominantly in the same regions.
[citation needed] The second theory comes from the idea that meiosis evolved from bacterial transformation, with the function of propagating genetic diversity.
[9][16] Thus, this evidence suggests that it is a question of whether cross over is linked to DNA repair or bacterial transformation, as the two do not appear to be mutually exclusive.
[22] Nevertheless, this mutant gave rise to spore viability patterns suggesting that segregation of non-exchange chromosomes occurred efficiently.
[23][24] The grasshopper Melanoplus femur-rubrum was exposed to an acute dose of X-rays during each individual stage of meiosis, and chiasma frequency was measured.
[25] Irradiation during the leptotene-zygotene stages of meiosis (that is, prior to the pachytene period in which crossover recombination occurs) was found to increase subsequent chiasma frequency.
Similarly, in the grasshopper Chorthippus brunneus, exposure to X-irradiation during the zygotene-early pachytene stages caused a significant increase in mean cell chiasma frequency.
These results suggest that X-rays induce DNA damages that are repaired by a crossover pathway leading to chiasma formation.
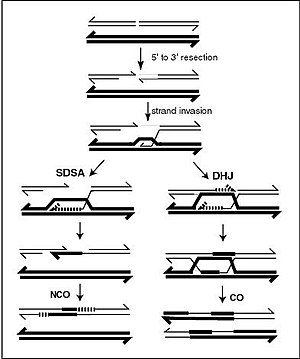
[citation needed] Double strand breaks (DSBs) are repaired by two pathways to generate crossovers in eukaryotes.
[28] Specifically, SLX4 knockout mice largely phenocopies the MUS81 knockout—once again, an elevated class I crossovers while normal chiasmata count.
An individual gamete inherits a complete haploid complement of alleles on chromosomes that are independently selected from each pair of chromatids lined up on the metaphase plate.
This leads to the notion of "genetic distance", which is a measure of recombination frequency averaged over a (suitably large) sample of pedigrees.
Linkage disequilibrium describes a situation in which some combinations of genes or genetic markers occur more or less frequently in a population than would be expected from their distances apart.
Because chromosomal regions composed of transposons have large quantities of identical, repetitious code in a condensed space, it is thought that transposon regions undergoing a crossover event are more prone to erroneous complementary match-up;[35] that is to say, a section of a chromosome containing a lot of identical sequences, should it undergo a crossover event, is less certain to match up with a perfectly homologous section of complementary code and more prone to binding with a section of code on a slightly different part of the chromosome.
[citation needed] While the motivating factors behind unequal recombination remain obscure, elements of the physical mechanism have been elucidated.