Clostridioides difficile toxin B
[4] Toxin B has four different structural domains: catalytic, cysteine protease, translocation, and receptor binding.
[6] The translocation region contains a hydrophobic stalk-like structure, which may help residues 958–1130 in forming membrane spanning pores.
Both cysteine proteases and a majority of translocation regions harbor hydrophobic proteins, which show access to TcdB and other toxins crossing the cell membranes.
the Rho family; Rac, and Cdc42 inside the target cells disturb signal transduction mechanisms, which leads to dysfunctioning of actin cytoskeleton, cell-cell junction, and apoptosis (Fig.
This phenomenon takes place by a binding receptor region, which enables toxin to enter host cells[citation needed].
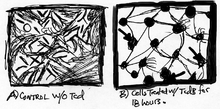
Figure 3 shows the probable effect of toxin B on a cell's surface; membrane blebbing (black arrows).
As a consequence, cell-cell junctions are disrupted, which enhances epithelial permeability of toxin B and fluid accumulation in the lumen.
[25] Several studies have indicated that Rho exhibits posttranslational modification through prenylation and carboxymethylation, which occurs in the cytoplasmic side of the plasma membrane, hence, the exchange of GTP to GDP.
[26] When TcdB binds to Rho and other small GTPases, GTP hydrolyzes to GDP, which leads to GTP-bound (active) to GDP-bound (inactive) (Fig.
[29] Shoshan et al., also showed that TcdB inhibited the receptor signaling activity by deactivating the Rho proteins via phospholipase D.[29] TcdB accesses the interior of the cell through clathrin-mediated endocytosis,[30] When toxin B is part of the cytosol, the glucosyltransferase passes through the endosomal membrane, which decreases pH, induces translocation and finally leads to morphological changes of translocation region residues (958–1130).
[31] When cells are infected with TcdB in an acidic environment, it attenuates toxins and causes shape rearrangements (Fig.
[31] As a consequence of acidic pH, TcdB displays clear differences in original fluorescence of tryptophan, susceptibility of proteases, and hydrophobic surfaces.
[31] Another group has shown that acidification leads to conformational changes of the toxin and, more importantly, helps to form pores.
[20] A majority of the studies used TcdB strain 630 to show the pore formation activity of C. difficile toxins.
[17] The encoding gene tcdE, located between tcdB and tcdA, is analogous to holin proteins, thus, it is suggested that tcdE works as a facilitator gene that enhances the release or secretion of TcdA and TcdB consequently increasing the permeability of the host cell membrane.
In order to detect the toxin B level in C. difficile, clinicians extensively use cell culture assays derived from stool specimens from patients with PMC.
[35][36] Moreover, knowing the role of antibiotics on the causes of PMC, many earlier studies focused on the effects of antimicrobials production of toxins.
As a result, studies were able to conclude that the subinhibitory nature of vancomycin and penicillin levels were increasing the toxin production in cultures of C.
Another study illustrated that the high levels of toxin production of TcdB were observed in complex mediums such as brain and heart infusion broth.
[40][41][42] Another study has shown that limiting biotin in well-characterized medium increases the production of TcdB by 64-fold and TcdA by 35-fold.
Clinical laboratories have identified these toxins in patients' stool based on antibody and cytotoxicity assays.
[9] Bezlotoxumab is a human monoclonal antibody designed for the prevention of recurrence of Clostridium difficile infections.
This has been deduced from in vivo experiments where toxin productions of TcdA were more severe than TcdB with antibiotics cecitis.
[38][46] Later, several studies showed that TcdB plays a major role in the disease of PMC and ADD.
[47] Furthermore, later studies have shown that a purified form of TcdB is a more lethal enterotoxin in comparison to TcdA, and also, that intestinal epithelium is severely damaged and causes an acute inflammatory response.
[48] With better understanding of the toxin, researchers were able to state that TcdB is the major virulence factor that causes CDI over TcdA.
However, when TcdA is present in the gut, it helps to facilitate TcdB's activity to have broader impacts, consequently, affecting multiple organ systems.