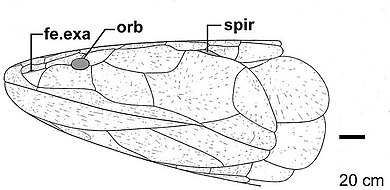
Eusthenodon
[7][4] The large size, predatory ecology, and evolutionarily derived characters possessed by Eusthenodon likely contributed to its ability to occupy and flourish in the numerous localities across the world mentioned above.
[3][8][9] As a result, it has been hypothesized that diversification of Eusthenodon and other morphologically similar tristichopterids were not restricted by biogeographical barriers and were instead limited only by their individual ecologies and mobility.
[5][9][8] Consequently, the majority of available literature covering Eusthenodon primarily focus on the intricacies of the bones associated with the skull in order to investigate the genus and while others draw conclusions from the known characters of Tristichopteridae.
[8] The further expansion of snout length in many tetrapod species may also be further evidence supporting the tendency of increasingly longer frontoethmoidal shields present in subsequent clades closely related to eusthenopterids including the late eopods.
[12][8] On the contrary, the pineal foramen is much smaller in size and is positioned distinctively posterior both to the center of radiation of the frontal and the postorbital bone of the frontoethmoidal shield.
[8] One theory to explain the trend observed in the posterior shift of the extratemporal bone in more derived fishes suggests that the change in head proportions contributed to a more streamlined body shape and enhanced its maneuverability and speed in its aquatic environment.
[8][4] In a study presented by Gael Clement in 2009, in which a newly discovered tristichopterid assemblage was described, it was found that the enlarged teeth were predominantly in line with the tooth row of the premaxilla and they did not occur in pairs.
[8][5] The presence of the enlarged pseudo fangs on the premaxilla in Eusthenodon, supported its phylogenetic position within the Tristichopteridae clade as similar dentition patterns are found in other closely related derived tristichopterids.
Additionally, the presence of a distinctive blade-like vertical lamina present on the anterior coronoid exists in most other tristichopterids but is absent in derived genera such as Eusthenodon.
[8] This reduction of marginal coronoid teeth supports the phylogenetic association of Eusthenodon, Mandageria, and Cabonnichthys and serves as a derived characteristic of late tristichopterids.
In line with the features described by Berg (1955) to be the significant diagnostic characters of Tristichopteridae, Eusthenodon possesses proportionately large, distinctively round scales without cosmine that exhibit a reticular pattern of ridges with rare appearance of independent tubercles.
Ahlberg (2008) after performing a maximum parsimony analysis of the interrelationships of within the clade:[9] Gogonasus Medoevia Tristichopterus Eusthenopteron Jarvikina Notorhizodon Platycephalichthys Cabonnichthys Mandageria Langlieria Eusthenodon