Exoelectrogen
Pelotomaculum thermopropioncum has been observed linked to Methanothermobacter thermautotrophicus by a pilus (external cell structures used in conjugation and adhesion) that was determined to be electrically conductive.
[1] In addition to S. oneidensis MR-1, exoelectrogenic activity has been observed in the following strains of bacteria without an exogenous mediator: Shewanella putrefaciens IR-1, Clostridium butyricum, Desulfuromonas acetoxidans, Geobacter metallireducens, Geobacter sulfurreducens, Rhodoferax ferrireducens, Aeromonas hydrophilia (A3), Pseudomonas aeruginosa, Desulfobulbus propionicus, Geopsychrobacter electrodiphilus, Geothrix fermentans, Shewanella oneidensis DSP10, Escherichia coli, Rhodopseudomonas palustris, Brucella anthropi YZ-1, Desulfovibrio desulfuricans, Acidiphilium sp.3.2Sup5, Klebsiella pneumoniae L17, Thermincola sp.strain JR, Pichia anomala.
Under low microbial population densities, usage of electron shuttles and chelators synthesized by the exoelectrogen may be energetically costly due to insufficient concentrations of such molecules required for recovery and reuse.
While these proteins are diverse (taking on both membrane-bound or soluble forms), their common locations in the outer membrane or periplasm in Gram-negative and Gram-positive bacteria provide intimate contact for electron transfer.
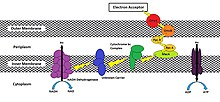
[4] As an example in Shewanella oneidensis MR-1, transport is characterized through a series of redox and structural proteins[11] extending from the cytoplasmic membrane to the outer cell surface (similar to Figure 1).
Flavins are secreted which are thought to bridge the “gap” between cell surface protein(s) and the external metal, which may alleviate the need for immediate contact and facilitate transfer at a distance.
Furthermore, G. sulferreducens produces electrically conductive pili (nanowires) with OmcS oxidoreductase enzymes embedded on its surface,[12] demonstrating the usage of multiple exoelectrogenic transfer methods.