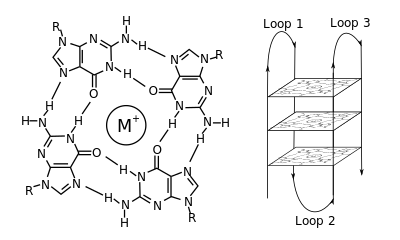
G-quadruplex
Short sequences, consisting of only a single contiguous run of three or more guanine bases, require four individual strands to form a quadruplex.
Analysis of human, chimpanzee, mouse and rat genomes showed enormous number of potential G-quadruplex (pG4)-forming sequences in non-telomeric regions.
[6] In addition, there was found more than one-billion-year conserved G-quadruplex locus in plants and algae, in gene encoding large subunit of RNA polymerase II.
Quadruplex formation may be potentially damaging for a cell; the helicases WRN and Bloom syndrome protein have a high affinity for resolving DNA G-quadruplexes.
[44] Function of G-quadruplexes have also been reported in allowing programmed recombination of immunologlobin heavy genes and the pilin antigenic variation system of the pathogenic Neisseria.
[54] The BER pathway is signalled when it indicates an oxidative DNA base damage, where structures like, 8-Oxoguanine-DNA glycosylase 1 (OGG1), APE1 and G-quadruplex play a huge role in its repair.
This form allows for the base excision repair (BER) enzyme OGG1 to bind and remove the oxidative damage with the help of APE1, resulting in an AP site.
[59] Upon insertion of 8-oxo-dG into thymidine kinase gene of humans, it was determine that if 8-oxo-dG was left unchecked and not repaired by BER, it can lead to frequent mutations and eventually carcinogenesis.
[65] Some of these oncogenes include c-KIT, PDGF-A, c-Myc and VEGF, showing the importance of this secondary structure in cancer growth and development.
[66] Current therapeutic research actively focuses on targeting this stabilization of G-quadruplex structures to arrest unregulated cell growth and division.
With this product, the c-Myc protein functions in the processes of apoptosis and cell growth or development and as a transcriptional control on human telomerase reverse transcriptase.
The formation of an intramolecular G-quadruplex structure has been shown through studies on the polypurine tract of the promoter region of the VEGF gene.
The guanine rich sequence in the promoter region for this pathway exudes a necessity for baseline transcription of this receptor tyrosine kinase.
[74] Ligand design and development remains an important field of research into therapeutic reagents due to the abundance of G-quadruplexes and their multiple conformational differences.
When bound, MM41's central chromophore is situated on top of the 3’ terminal G-quartet and the side chains of the ligand associate to the loops of the quadruplex.
Generally, a simple pattern match is used for searching for possible intrastrand quadruplex forming sequences: d(G3+N1-7G3+N1-7G3+N1-7G3+), where N is any nucleotide base (including guanine).
In one study,[82] it was found that the observed number per base pair (i.e. the frequency) of these motifs has increased rapidly in the eumetazoa for which complete genomic sequences are available.
This suggests that the sequences may be under positive selection enabled by the evolution of systems capable of suppressing non-B structure formation.
[87] The topology of the G-quadruplex structure can be determined by monitoring the positive or negative circular dichroism (CD) signals at specific wavelengths.
As the quantum yields of the intrinsic G-quadruplex fluorescence are higher compared to those of the constitutive single strands, their formation[90] or their melting[91] can be detected by monitoring their emission.
This repeat expansion promotes DNA methylation and other epigenetic heterochromatin modifications of FMR1 that prevent the transcription of the gene, leading to pathological low levels of FMRP.
[106][107] Antisense-mediated interventions and small-molecule ligands are common strategies used to target neurological diseases linked to G-quadruplex expansion repeats.
[108] Antisense therapy is the process by which synthesized strands of nucleic acids are used to bind directly and specifically to the mRNA produced by a certain gene, which will inactivate it.
Antisense oligonucleotides (ASOs) are commonly used to target C9orf72 RNA of the G-quadruplex GGGGCC expansion repeat region, which has lowered the toxicity in cellular models of C9orf72.
[109][110][111] ASOs have previously been used to restore normal phenotypes in other neurological diseases that have gain-of-function mechanisms, the only difference is that it was used in the absence of G-quadruplex expansion repeat regions.
These decoys are typically composed of a G-rich sequence that can form a stable G-quadruplex structure and a short linker region that can be modified to optimize their properties.
[116] When introduced to cancer cells the decoy can intercept associated transcription factors and bind them leading to the regulation of gene expression.
Decoys have been successfully demonstrated to inhibit oncogenic KRAS in SCID mice leading to reduced tumour growth and increased median survival time.
A disadvantage of using small-molecule ligands as a therapeutic technique is that specificity is difficult to manage due to the variability of G-quadruplexes in their primary sequences, orientation, thermodynamic stability, and nucleic acid strand stoichiometry.
[120] Small-molecule ligands, composed primarily of lead, can target GGGGCC repeat regions as well and ultimately decreased both repeat-associated non-ATG translation and RNA foci in neuron cells derived from patients with Amyotrophic lateral sclerosis (ALS).