Garudimimus
The genus is known from a single specimen found in 1981 by a Soviet-Mongolian paleontological expedition in the Bayan Shireh Formation and formally described in the same year by Rinchen Barsbold; the only species is Garudimimus brevipes.
It was previously thought that this primitive ornithomimosaurian possessed a lacrimal "horn" at the top of the skull, in front of the eye socket.
However, with the description of new specimens of Deinocheirus in 2014, it was found that the latter was the sister taxon of Garudimimus, grouping within the Deinocheiridae—ornithomimosaurs not adapted for running or agile movements.
The pelvic girdle and hindlimbs of Garudimimus show that the musculature of the legs was not as well-developed as in the fast-running ornithomimids, therefore, indicating poor cursorial capacities.
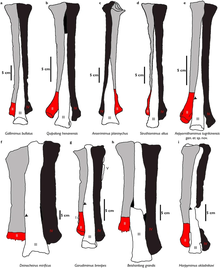
In 1981, during a Soviet-Mongolian paleontological expedition to the Gobi Desert, a relatively small theropod skeleton was discovered by the team at Bayshi Tsav in the Late Cretaceous Bayan Shireh Formation, Southeastern Mongolia.
The remains are catalogued as MPC-D 100/13 (Mongolian Palaeontological Center; originally GIN 100/13) and represent a rather complete and articulated skeleton.
In the same year this specimen was formally and briefly described by the Mongolian paleontologist Rinchen Barsbold as the holotype of the genus and species Garudimimus brevipes.
The generic name, Garudimimus, combines a reference to Garuda which are legendary winged creatures from Mongolian Buddhist mythology and Latin mimus (meaning mimic).
[1][4] In 1988 the North American author Gregory S. Paul illustrated Garudimimus with a prominent nasal horn unlike any other ornithomimosaur, considering this feature as actually preserved.
[5] Another interpretation was made also in 1988 by Philip J. Currie and Dale Alan Russell who referred the metatarsus of the holotype to Oviraptor sp.
The supposed orbital horn is actually the disarticulated left prefrontal bone and verified that the metatarsus did not suffer taphonomical (changes during decay and fossilisation) distortion and is non-arctometatarsalian.
[4] Although Garudimimus is solely known from MPC-D 100/13, additional specimens may potentially be present among a large bonebed composed of at least five individuals in Bayshi Tsav and several undescribed ornithomimosaurs from other localities in the Bayan Shireh Formation.
[4] The edentulous condition of both premaxilla and dentary with the addition of numerous foramina in their surfaces, indicate that a prominent rhamphotheca (horny beak) was present in life.
The jugal had a triangular main body with very thin-walled processes (bony extensions); it also gave form to the lower border of the eye socket.
[4] Putative large foramen and deep fossae (depressions) were present on the anterior neural arches, which may indicate caudal pneumaticity in Garudimimus.
Kobayashi and Barsbold noted that this taxon shared traits with primitive and derived ornithomimosaurs such as four-toed feet and toothless jaws.
Lee and team performed a large phylogenetic analysis for the Deinocheiridae and the three members were found in this grouping: Beishanlong, Garudimimus and Deinocheirus.
[4][22] In 2011, Lars Schmitz and Ryosuke Motani measured the dimensions of the sclerotic ring and eye socket in fossil specimens of dinosaurs and pterosaurs, as well as some living species.
Schmitz and Motani separated ecological and phylogenetic factors and by examining 164 living species, they noticed that eye measurements are quite accurate when inferring diurnality, cathemerality, or nocturnality in tetrapods.
[25] However, this analysis was criticized in the same year by Hall and colleagues arguing against the methods employed by Schmitz and Motani in that there is a considerable degree of similarities on scleral anatomy between diurnal, cathemeral and nocturnal animals.
[26] Schmitz and Motani made a response to this comment affirming their research and methodology properly categorized extinct dinosaur behaviors based on extant species.
[28] Kobayashi and Barsbold stated in 2005 that the absence of dentition on the lower jaw and the recurved shape of the same indicates that Garudimimus was fairly more similar to ornithomimids when consuming food than to other primitive ornithomimosaurs.
[29][25] In 2015, Andrew R. Cuff and Emily J. Rayfield performed CT scans on taphonomically distorted specimen skulls of Garudimimus, Ornithomimus and Struthiomimus in order to be retrodeformed—a method used to produce the original state of a deformed object.
The scans and relatively completeness of the specimens allowed the reconstruction of the adductor (motion-inducing) musculature of the lower jaw, which was then mapped onto the skulls.
Cuff and Rayfield pointed out that the bite force of Garudimimus may be limited for having to use the mandible of Struthiomimus in the reconstructions or that MPC-D 100/13 is a sub-adult individual.
However, Button and Zanno pointed out that the relationship between herbivory and size is not always related and there is no lineal trend of increasing mass in ornithomimosaurs.
Nabavizadeh noted that ornithomimosaurs were orthal feeders and raised their jaws isognathously whereby the upper and lower teeth of each side occluded (contacted each other) at once.
[35] Numerous scale-like cross-stratifications at the Baynshire locality are indicators of large meanders, and the vast majority of these water bodies may have drained the eastern part of the Gobi Desert.
[36][37] Based on the similar fossil assemblages, a correlation between the Bayan Shireh and Iren Dabasu formations has been proposed by numerous authors.
[44] Several species were separated by niche differentiation, such as Erlikosaurus and the large Segnosaurus,[43] or the grazer Talarurus and browser Tsagantegia, another ankylosaurid from the formation.