Deinocheirus
After more complete remains were found, Deinocheirus was shown to be a primitive ornithomimosaurian, most closely related to the smaller genera Garudimimus and Beishanlong, together forming the family Deinocheiridae.
The first known fossil remains of Deinocheirus were discovered by Polish palaeontologist Zofia Kielan-Jaworowska on July 9, 1965, at the Altan Ula III site (coordinates: 43°33.987′N 100°28.959′E / 43.566450°N 100.482650°E / 43.566450; 100.482650) in the Nemegt Basin of the Gobi Desert.
[1] The paucity of known Deinocheirus remains inhibited a thorough understanding of the animal for almost half a century onwards, and the scientific literature often described it as among the most "enigmatic", "mysterious", and "bizarre" of dinosaurs.
[7] In 2012, Phil R. Bell, Philip J. Currie, and Yuong-Nam Lee announced the discovery of additional elements of the holotype specimen, including fragments of gastralia, found by a Korean-Mongolian team which re-located the original quarry in 2008.
[6] In 2013, the discovery of two new Deinocheirus specimens were announced before the annual Society of Vertebrate Paleontology (SVP) conference by Lee, Barsbold, Currie, and colleagues.
MPC-D 100/128, a subadult specimen, was found by scientists in the Altan Ula IV locality (coordinates: 43°36.091′N 100°27.066′E / 43.601517°N 100.451100°E / 43.601517; 100.451100) of the Nemegt Formation during the Korea-Mongolia International Dinosaur Expedition in 2006, but had already been damaged by fossil poachers.
[9] Currie stated in an interview that it was a policy of their team to investigate quarries after they had been looted and recover anything of significance, and that finding any new Deinocheirus fossils was cause for celebration, even without the poached parts.
[10] The poached elements were spotted in a private European collection by the French fossil trader François Escuillé, who notified Belgian palaeontologist Pascal Godefroit about them in 2011.
[11] The recovered material consisted of a skull, a left hand, and feet, which had been collected in Mongolia, sold to a Japanese buyer, and resold to a German party (the fossils also passed through China and France).
American palaeontologist Thomas R. Holtz stated in an interview that the new Deinocheirus remains looked like the "product of a secret love affair between a hadrosaur and Gallimimus".
[14] By 2017, the Mongolian government had increased its effort to seize poached fossils from collectors and repatriate them, but proving their provenance had become a scientific and political concern.
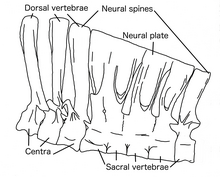
The neural spines had a system of interconnecting ligaments, which stiffened the vertebral column allowing it to support the abdomen while transmitting the stress to the hips and hindlimbs.
A 2006 study by Kobayashi and Barsbold found Deinocheirus to be possibly the most primitive ornithomimosaur, but was unable to further resolve its affinities, due to the lack of skull and hindlimb elements.
[5] A cladistic analysis accompanying the 2014 description of the two much more complete specimens found that Deinocheirus formed a clade with Garudimimus and Beishanlong, which were therefore included in the Deinocheiridae.
[26] The blunt and short hand-claws of Deinocheirus were similar to those of the therizinosaur Alxasaurus, which indicates the long arms and claws were used for digging and gathering plants.
The brain was proportionally small and compact, and its reptile encephalization quotient (brain-body ratio) was estimated at 0.69, which is low for theropods, and similar to sauropods.
It has been proposed that pneumatisation was used to reduce the mass of large bones (associated with gigantic size in the case of Deinocheirus), that it was related to high metabolism, balance during locomotion, or used for thermoregulation.
[24] A bone microstructure study presented at the European Association of Vertebrate Palaeontologists in 2015 showed that Deinocheirus probably had a high metabolic rate, and grew rapidly before reaching sexual maturity.
[28] A histological study of a gastralia fragment from the holotype presented at a 2018 conference showed that its internal structure was similar to that of ossified tendons of other theropods.
The attachment sites for the muscles that open and close the jaws were very small in comparison to the size of the skull, which indicates Deinocheirus had a weak bite force.
The depth of the lower jaw indicates the presence of a large tongue, which could have assisted the animal in sucking in food material obtained with the broad beak when foraging on the bottom of freshwater bodies.
[30] A 2022 article by Waisum Ma and colleagues examined how feeding mechanics varied between different non-bird coelurosaurian groups through finite element analysis, revealing that they all underwent reduction of feeding-related stress in their jaws.
[33] Osmólska and Roniewicz reported palaeopathologies in the holotype specimen such as abnormal pits, grooves and tubercles on the first and second phalanx of the left second finger that may have been the result of injuries to the joint between the two bones.
This geologic formation has never been dated radiometrically, but the fauna present in the fossil record indicate it was probably deposited during the early Maastrichtian age, at the end of the Late Cretaceous about 70 million years ago.
Such large river channels and soil deposits are evidence of a far more humid climate than those found in the older Barun Goyot and Djadochta formations.
Along with Deinocheirus, the discoveries of Therizinosaurus and Gigantoraptor show that three groups of herbivorous theropods (ornithomimosaurs, therizinosaurs and oviraptorosaurs), independently reached their maximum sizes in the late Cretaceous of Asia.
[35][39][40] Mammal fossils are rare in the Nemegt Formation, but many birds have been found, including the enantiornithine Gurilynia, the hesperornithiform Judinornis, as well as Teviornis, a possible Anseriform.
Herbivorous dinosaurs of the Nemegt Formation include ankylosaurids such as Tarchia, the pachycephalosaurian Prenocephale, large hadrosaurids such as Saurolophus and Barsboldia, and sauropods such as Nemegtosaurus, and Opisthocoelicaudia.
[35][41] Predatory theropods that lived alongside Deinocheirus include tyrannosauroids such as Tarbosaurus, Alioramus, and Bagaraatan, and troodontids such as Borogovia, Tochisaurus, and Zanabazar.
Theropod groups with both omnivorous and herbivorous members include therizinosaurs, such as Therizinosaurus, oviraptorosaurians, such as Elmisaurus, Nemegtomaia, and Rinchenia, and other ornithomimosaurians, such as Anserimimus and Gallimimus.