Gas exchange
Gases are constantly consumed and produced by cellular and metabolic reactions in most living things, so an efficient system for gas exchange between, ultimately, the interior of the cell(s) and the external environment is required.
Some small multicellular organisms, such as flatworms, are also able to perform sufficient gas exchange across the skin or cuticle that surrounds their bodies.
However, in most larger organisms, which have small surface-area to volume ratios, specialised structures with convoluted surfaces such as gills, pulmonary alveoli and spongy mesophylls provide the large area needed for effective gas exchange.
In aerobic organisms, gas exchange is particularly important for respiration, which involves the uptake of oxygen (O2) and release of carbon dioxide (CO2).
Other gas-exchange processes are important in less familiar organisms: e.g. carbon dioxide, methane and hydrogen are exchanged across the cell membrane of methanogenic archaea.
In comparison to this small volume, the surface area of its cell membrane is very large, and adequate for its gas-exchange needs without further modification.
Sponges do not require a circulatory system or specialised gas exchange organs, because their feeding strategy involves one-way pumping of water through their porous bodies using flagellated collar cells.
A countercurrent system such as this maintains a steep concentration gradient along the length of the gas-exchange surface (see lower diagram in Fig.
[9] The gas-containing environmental water is drawn unidirectionally across the gas-exchange surface, with the blood-flow in the gill capillaries beneath flowing in the opposite direction.
This means the magnitude of the gradient is variable along the length of the gas-exchange surface, and the exchange will eventually stop when an equilibrium has been reached (see upper diagram in Fig.
The membrane across which gas exchange takes place in the alveoli (i.e. the blood-air barrier) is extremely thin (in humans, on average, 2.2 μm thick).
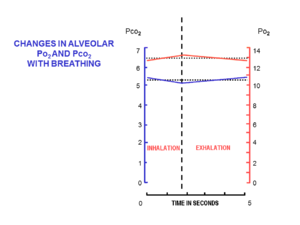
This relatively large volume of air that is semi-permanently present in the alveoli throughout the breathing cycle is known as the functional residual capacity (FRC).
[15] The entry of such a small volume of fresh air with each inhalation, ensures that the composition of the FRC hardly changes during the breathing cycle (Fig.
This ensures that the partial pressures of oxygen and carbon dioxide in the blood leaving the alveolar capillaries, and ultimately circulates throughout the body, are the same as those in the FRC.
This anatomy, and the fact that the lungs are not emptied and re-inflated with each breath, provides mammals with a "portable atmosphere", whose composition differs significantly from the present-day ambient air.
[18] The composition of the air in the FRC is carefully monitored, by measuring the partial pressures of oxygen and carbon dioxide in the arterial blood.
On its passage through the lungs the blood comes into close contact with the alveolar air, separated from it by a very thin diffusion membrane which is only, on average, about 2 μm thick.
This is very tightly controlled by the continuous monitoring of the arterial blood gas tensions (which accurately reflect partial pressures of the respiratory gases in the alveolar air) by the aortic bodies, the carotid bodies, and the blood gas and pH sensor on the anterior surface of the medulla oblongata in the brain.
In the long run these can be compensated by renal adjustments to the H+ and HCO3− concentrations in the plasma; but since this takes time, the hyperventilation syndrome can, for instance, occur when agitation or anxiety cause a person to breathe fast and deeply[20] thus blowing off too much CO2 from the blood into the outside air, precipitating a set of distressing symptoms which result from an excessively high pH of the extracellular fluids.
The lamellae contain capillaries that provide a large surface area and short diffusion distances, as their walls are extremely thin.
This countercurrent maintains steep concentration gradients along the entire length of each capillary (see the diagram in the "Interaction with circulatory systems" section above).
[9] Amphibians have three main organs involved in gas exchange: the lungs, the skin, and the gills, which can be used singly or in a variety of different combinations.
To increase the rate of gas exchange by diffusion, amphibians maintain the concentration gradient across the respiratory surface using a process called buccal pumping.
[13] Selective bronchoconstriction at the various bronchial branch points ensures that the air does not ebb and flow through the bronchi during inhalation and exhalation, as it does in mammals, but follows the paths described above.
At night, plants respire, and gas exchange partly reverses: water vapor is still lost (but to a smaller extent), but oxygen is now taken up and carbon dioxide released.
If the environmental conditions (humidity, CO2 concentration, light and temperature) are fully controlled, the measurements of CO2 uptake and water release reveal important information about the CO2 assimilation and transpiration rates.
[40][41] Simpler methods can be used in specific circumstances: hydrogencarbonate indicator can be used to monitor the consumption of CO2 in a solution containing a single plant leaf at different levels of light intensity,[42] and oxygen generation by the pondweed Elodea can be measured by simply collecting the gas in a submerged test-tube containing a small piece of the plant.
[citation needed] The roundworms (Nematoda), flatworms (Platyhelminthes), and many other small invertebrate animals living in aquatic or otherwise wet habitats do not have a dedicated gas-exchange surface or circulatory system.
[citation needed] Other aquatic invertebrates such as most molluscs (Mollusca) and larger crustaceans (Crustacea) such as lobsters, have gills analogous to those of fish, which operate in a similar way.
[48] The other main group of terrestrial arthropod, the arachnids (spiders, scorpion, mites, and their relatives) typically perform gas exchange with a book lung.