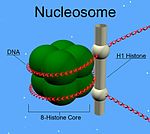
Histone H1
Metazoan H1 proteins feature a central globular "winged helix" domain and long C- and short N-terminal tails.
Ionic extraction of linker histones from native or reconstituted chromatin promotes its unfolding under hypotonic conditions from fibers of 30 nm width to beads-on-a-string nucleosome arrays.
Cellular studies have shown that overexpression of H1 can cause aberrant nuclear morphology and chromatin structure, and that H1 can serve as both a positive and negative regulator of transcription, depending on the gene.
[18][19] Complete knockout of H1 in vivo has not been achieved in multicellular organisms due to the existence of multiple isoforms that may be present in several gene clusters, but various linker histone isoforms have been depleted to varying degrees in Tetrahymena, C. elegans, Arabidopsis, fruit fly, and mouse, resulting in various organism-specific defects in nuclear morphology, chromatin structure, DNA methylation, and/or specific gene expression.
[19] The H1 family in animals includes multiple H1 isoforms that can be expressed in different or overlapping tissues and developmental stages within a single organism.
The reason for these multiple isoforms remains unclear, but both their evolutionary conservation from sea urchin to humans as well as significant differences in their amino acid sequences suggest that they are not functionally equivalent.
[3][30][31][32] Despite having more negative charges than somatic isoforms, H1M binds with higher affinity to mitotic chromosomes in Xenopus egg extracts.