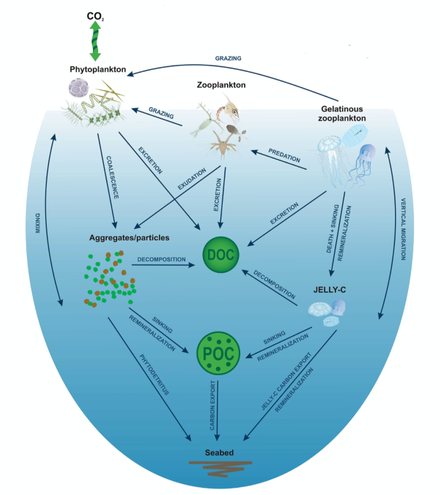
Gelatinous zooplankton
Jellyfish, and more gelatinous zooplankton in general, which include salps and ctenophores, are very diverse, fragile with no hard parts, difficult to see and monitor, subject to rapid population swings and often live inconveniently far from shore or deep in the ocean.
[7] But jellyfish bloom in vast numbers, and it has been shown they form major components in the diets of tuna, spearfish and swordfish as well as various birds and invertebrates such as octopus, sea cucumbers, crabs and amphipods.
Traditionally gelatinous predators were thought ineffectual providers of marine trophic pathways, but they appear to have substantial and integral roles in deep pelagic food webs.
They also can rapidly reproduce on a time scale of days and, under favorable environmental conditions, some species form dense blooms that extend for many square kilometers.
Globally, gelatinous zooplankton abundance and distribution patterns largely follow those of temperature and dissolved oxygen as well as primary production as the carbon source.
[21] However, gelatinous zooplankton cope with a wide spectrum of environmental conditions, indicating the ability to adapt and occupy most available ecological niches in a water mass.
In terms of Longhurst regions (biogeographical provinces that partition the pelagic environment,[28][29] the highest densities of gelatinous zooplankton occur in coastal waters of the Humboldt Current, NE U.S.
[32][11] Large amounts of jelly carbon are quickly transferred to and remineralized on the seabed in coastal areas, including estuaries, lagoons and subtidal/intertidal zones,[15] shelves and slopes,[33][34][35] the deepsea.
[41][42] Despite the high lability of jelly‐C,[43][39] a remarkably large amount of biomass arrives at the seabed below 1,000 m. During sinking, jelly‐C biochemical composition changes via shifts in C:N:P ratios as observed in experimental studies.
Furthermore, jelly‐C deposits tend not to build up at the seafloor over a long time, such as phytodetritus (Beaulieu, 2002), being consumed rapidly by demersal and benthic organisms [41] or decomposed by microbes.
[58] In 2013, Lebrato et al. determined the average sinking speed of jelly‐C using Cnidaria, Ctenophora, and Thaliacea samples, which ranged from 800 to 1,500 m day−1 (salps: 800–1,200 m day−1; scyphozoans: 1,000–1,100 m d−1; ctenophores: 1,200–1,500 m day−1; pyrosomes: 1,300 m day−1).