KcsA potassium channel
The pH activated protein possesses two transmembrane segments and a highly selective pore region, responsible for the gating and shuttling of K+ ions out of the cell.
[6] Three years later, a higher resolution model was produced by Morais-Cabral and Zhou after monoclonal Fab fragments were attached to KcsA crystals to further stabilize the channel.
[7] In the early 2000s, evidence for the occupation of the selectivity filter by two K+ atom during the transport process emerged, based on energy and electrostatic calculations made to model the pore region.
In 2011, the crystal structure of full length KcsA was resolved to reveal that hindrance by the previously truncated residues permits only straightforward expansion of the intercellular ion passage region of the protein.
The structure of KcsA is that of an inverted cone, with a central pore running down the center made up of two transmembrane helices (the outer-helix M1 and the inner-helix M2), which span the lipid bilayer.
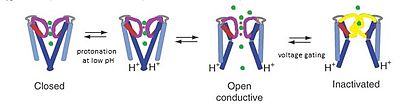
[16] In the nonconducting conformation of KcsA at pH 7, K+ is bound tightly to coordinating oxygens of the selectivity filter and the four TM2 helices converge near the cytoplasmic junction to block the passage of any potassium ions.
[5] The glycines and threonine coordinate with the K+ ion, while the side-chains of valine and tyrosine are directed into the protein core to impose geometric constraint on the filter.
As a result, the KcsA tetramer harbors four equal spaced K+ binding sites, with each side composed of a cage formed by eight oxygen atoms that sit on the vertices of a cube.
NMR studies performed by the Riek group show that pH sensitivity occurs in both the C-terminal TM2 region of the protein as well as with Tyr78 and Gly79 residues in the selectivity filter.
Exchanging negatively charged amino acids for neutral ones made the KcsA channel insensitive to pH even though there were no amino-acid changes at the transmembrane region.
NMR studies also suggest that a complex hydrogen bond network between Tyr78, Gly79, Glu71 and Asp80 exists in the KcsA filter region, and further acts as a pH-sensitive trigger for conductance.
After adding a depolarizing current to the channel, the reorientation of Glu71 towards the intracellular pore occurs, thereby disrupting the Glu71-Asp80 carboxyl-carboxylate pair that initially stabilizes the selectivity filter.
The orientation of amino acids in the filter region might play significant physiological role in modulating potassium fluxes in eukaryotes and prokaryotes under steady-state conditions.
Such tests can screen for drug-hERG channel interactions that cause acquired long QT syndrome, are essential for determining the cardiac safety of new medications.
Such models reveal the flexibility of the hERG channel and can consistently predict the binding affinity of a set of diverse ion channel-interacting ligands.