Ketoacyl synthase
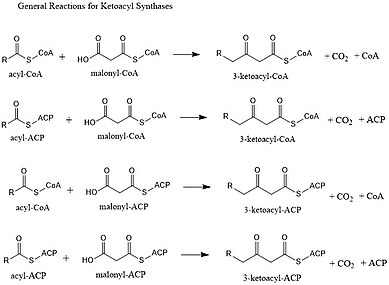
This reaction is a key step in the fatty acid synthesis cycle, as the resulting acyl chain is two carbon atoms longer than before.
They contain discrete functional domains responsible for specific catalytic activities of the reaction sequence, either on a single polypeptide chain or on two different multifunctional proteins.
Type II FASs are dissociated systems, meaning the component enzymes are independent proteins encoded by a series of separate genes.
[2] Polyketide synthases (PKS) are structurally and functionally related to FAS’s, both which are enzymes that catalyze the condensation of activated primary metabolites such as acetyl-CoA and malonyl-CoA.
They also both used a ketoreductase, dehydratase, and enoyl reductase to create a fully saturated acyl backbone.
Type III PKSs are multifunctional when choosing a starting unit, assembling the chain, and promoting the folding.
[5][1] The characteristic reaction of β-ketoacyl-ACP synthase III is malonyl-ACP + acetyl-CoA => acetoacyl-ACP + CO2 + CoA.
Cysteine, histidine, and asparagine form the catalytic triad in KAS III, which uses the ping-pong kinetic mechanism.
[1] In Escherichia coli, one organism KAS III is typically found in, KASIII is weakly inhibited by thiolactomycin.
[1] 3-Ketoacyl-CoA synthase I in Arabidopsis thaliana is involved in very long chain fatty acid synthesis, which plays a role in wax biosynthesis.
KS3 enzymes are primarily produced in bacteria, with a small number of eukaryotes and archaea.
It uses fatty acyl thioesters of ACP and CoA as substrates and has a specificity close to that of beta-ketoacyl-ACP synthase II.
For example, in certain cases, acyl-CoA itself inhibits the enzyme at high concentrations in Escherichia coli.
Cerulenin is known to inhibit synthase I in Carthamus tinctorius, Spinacia oleracea, Brassica napus, Allium ampeloprasu, Streptococcus pneumoniae, Escherichia coli, Mycobacterium tuberculosis, and many more.
[15] The reaction proceeds as such: In Streptococcus pneumoniae, for example, synthase II is used as an elongation condensing enzyme.
His 337 is also used for the condensation activity, specifically the stabilization of the negative charge on the malonyl thioester carbonyl in the transition state.
His303 is used to accelerate catalysis by deprotonating the water molecule to allow for a nucleophilic attack on malonate, thereby releasing bicarbonate.
For example, cerulenin inhibits synthase II in Spinacia oleracea, Allium ampelprasum, Escherichia coli, and Streptoccoccus pneumonia.
[17] A number of inhibitors include cerulenin in Sinapis alba, Daucus carota, and Phaseolus vulgaris, apigenin in Secale cereale and Avena sativa, and eriodictyol in Decale cereal, Daucus carota, and Xanthisma gracile.