mRNA surveillance
These mechanisms function at various steps of the mRNA biogenesis pathway to detect and degrade transcripts that have not properly been processed.
The translation of messenger RNA transcripts into proteins is a vital part of the central dogma of molecular biology.
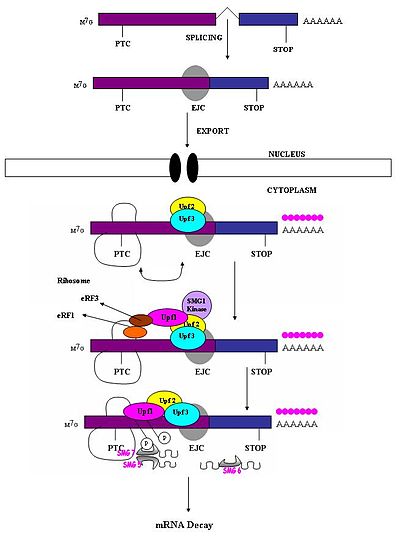
[4] Fidelity checks of mRNA molecules in the nucleus results in the degradation of improperly processed transcripts before export into the cytoplasm.
Cytoplasmic surveillance mechanisms assess mRNA transcripts for the absence of or presence of premature stop codons.
[5][6] Failure to recognize and decay these mRNA transcripts can result in the production of truncated proteins which may be harmful to the organism.
By causing decay of C-terminally truncated polypeptides, the NMD mechanism can protect cells against deleterious dominant-negative, and gain of function effects.
[7] PTCs have been implicated in approximately 30% of all inherited diseases; as such, the NMD pathway plays a vital role in assuring overall survival and fitness of an organism.
It has been observed that the ability of a nonsense codon to cause mRNA degradation depends on its relative location to the downstream sequence element and associated proteins.
[18] Furthermore, it has been observed that NMD mechanisms are not activated by nonsense transcripts that are generated from genes that naturally do not contain introns (e.g., Histone H4, Hsp70, melanocortin-4-receptor).
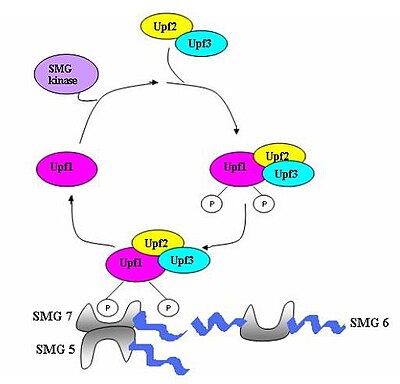
[7] When the ribosome reaches a PTC the translation factors eRF1 and eRF3 interact with retained EJC complexes though a multiprotein bridge.
[11] A second model proposes that a widely present feature in mRNA, such as a 3' poly-A tail, might provide the positional information required for recognition.
[17][25] In general, these mRNA molecules possess the stop codon very early in the reading frame (i.e. the PTC is AUG-proximal).
β-globulin mRNAs containing a nonsense mutation early in the first exon of the gene are more stable than NMD sensitive mRNA molecules.
[27] It has been demonstrated in other studies that the presence of this protein near AUG-proximal PTCs appears to promote the stability of these otherwise NMD sensitive mRNAs.
[29][30] These mRNA transcripts can arise from many different mechanisms such as premature 3' adenylation or cryptic polyadenylation signals within the coding region of a gene.
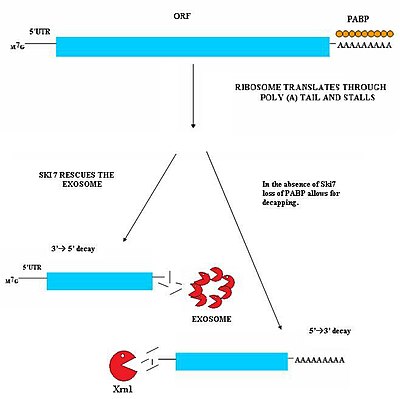
In this mechanism, the absence of Ski7 results in the loss of poly-A tail binding PABP proteins by the action of the translation ribosome.
While authentic targets of NGD are poorly understood, they appear to consist largely of mRNA transcripts on which ribosomes have stalled during translation.
This stall can be caused by a variety of factors including strong secondary structures, which may physically block the translational machinery from moving down the transcript.
[34] In some cases, the transcript is also cleaved in an endonucleolytic fashion near the stall site; however the identity of the responsible endonuclease remains contentious.